

5.2. Первинна структура нуклеїнових кислот
Нуклеотиди, що входять до складу нуклеїнових кислот, поєднані між собою за допомогою 3,5-фосфодіетерного зв’язку (рис. 5.8). Послідовність нуклеотидів визначає первинну структуру нуклеїнових кислот. Отже, утворюються два класи нуклеїнових кислот: ДНК і РНК. Як правило, генетичну інформацію несе ДНК. У деяких вірусів цю роль виконує РНК.
ДНК і РНК являють собою лінійні полімери. Майже всі РНК, за виключенням деяких РНК вірусної природи, є одноланцюговими, тоді як ДНК практично всі є дволанцюговими, побудованими за принципом комплементарності. Це накладає певні обмеження на нуклеотидний склад ДНК. Цю закономірність відкрито й описано Е. Чаргафором, і відома вона як правило Чаргаффа:
мольна частка аденіну дорівнює мольній частці тиміну, тобто А/Т = 1;
мольна частка гуаніну дорівнює мольній частці цитозину, тобто Г/Ц = 1.
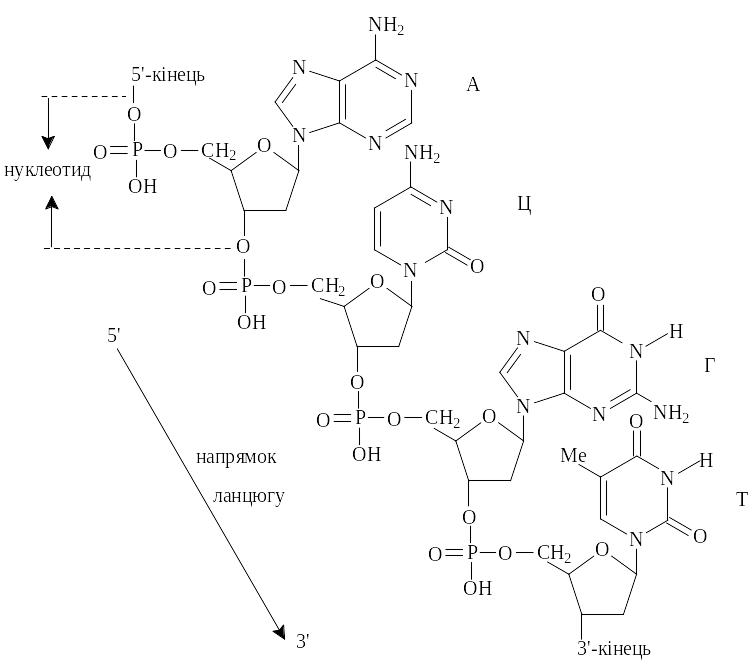
Рис.5.8. Схема будови нуклеїнових кислот. Ме = СН3
Отже, для кількісного визначення нуклеотидного складу подвійної спіралі ДНК достатньо знати одну змінну – мольну частку одного із чотирьох нуклеотидів. Нуклеотидний склад природних ДНК суттєво відрізняється. Так, у ДНК деяких бактерій мольна частка Г-Ц пар становить 75 %. У тварин цей параметр лежить у межах 40–50 %. Довжина ланцюгів рибонуклеїнових кислот не перевищує частки міліметра. Найкоротші з них тРНК, ступінь полімеризації яких становить приблизно 80. Найдовші РНК, це так звані гетерогенні ядерні РНК (гяРНК), містять до 2 105 нуклеотидів. Довжина ДНК коливається в більш широких межах, від частки міліметра (кілька тисяч нуклеотидів) до сантиметрів (≈109 нуклеотидів). Наприклад, молекули ДНК хромосоми людини містять від 48 млн до 240 млн пар основ і мають довжину від 1,6 до 8,2 см.
Процедура виділення високомолекулярних ДНК у чистому вигляді непроста, оскільки навіть витікання розчину ДНК із піпетки приводить до розриву ковалентних зв’язків під дією ваги, тобто до фрагментації.
У нуклеїнових кислотах виявлено різноманітні типи структур. Деякі з них є тимчасовими, тобто виникають при здійсненні нуклеїновими кислотами своїх функцій. На словах їх можна описати як одноланцюгові, дволанцюгові, дволанцюгові з одноланцюговими кінцями, розгалужені, циклічні одноланцюгові, циклічні дволанцюгові, структури типу катенанів.
Визначити нуклеотидну послідовність у ДНК довгий час не вдавалось. За останні 20 років у цій галузі відбувся величезний прогрес. На сьогодні розшифровано первинну структуру ДНК геному людини. Це зумовлено, головним чином, двома факторами. По-перше, було розроблено електрофоретичні методи розділення полінуклеотидів довжиною до 700 мономерів з точністю до одного нуклеотиду. По-друге, було відкрито ферменти – рестриктази, які розщеплюють довгі молекули ДНК за певними нуклеотидними послідовностями, що дозволяє отримувати однакові фрагменти ДНК.
5.3. Вторинна структура днк
Вторинну структуру ДНК запропоновано Дж. Уотсоном і Ф. Кріком у 1953 р. (Нобелівська премія в 1962 р.) на базі узагальнення цілої низки даних вивчення нуклеїнових кислот. Найважливіші з них: правила Чаргафа; 3,5-фосфодіетерний зв’язок між нуклеотидами; рентгенографічні дані препаратів ДНК, отримані М. Уілкінсом і Р. Франклін; -конфігурація глікозидного зв’язку; циклічна форма дезоксирибози.
Уотсон і Крік врахували розміри кожного нуклеотиду й побудували модель ДНК у вигляді подвійної спіралі й висунули ідею про специфічну взаємодію між комплементарними парами А і Т та Г і Ц. Характерною особливістю цих структур є однакова відстань С1- С1 у кожної пари, що забезпечує утворення подвійної спіралі однакового діаметра незалежно від нуклеотидного складу (рис. 5.9)
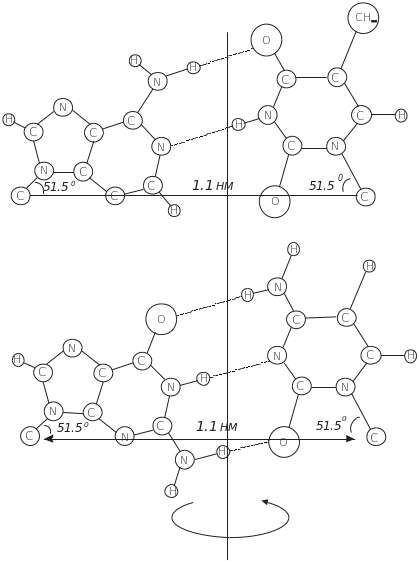
Рис. 5.9. Будова комплементарних пар нуклеозидів, що формують подвійну спіраль ДНК
Основи, що утворюють комплементарну пару, не лежать в одній площині, а повернуті відносно одна одної таким чином, що мають вигляд пропелера, кут якого визначається двогранним кутом між площинами основ.
Вирішальне значення для побудови просторової структури ДНК мали рентгенографічні дослідження, виконані Уілкінсом і Франклін. Вони отримали рентгенограми орієнтованих ниток ДНК (рис. 5.10).
Положення дифракційних максимумів (рефлексів) на рентгенограмі відповідає умові Вульфа–Брегга:
2b sin = n, (5.2)
де b – постійні гратки (відстань між площинами); – кут дифракції; n = 0,1, 2, 3… – порядок дифракції; - довжина хвилі. За формулою (5.2) великим значенням b відповідають малі кути .
На рентгенограмі В-форми ДНК можна виділити дві постійні гратки, які характеризують впорядковані структури, що розміщуються перпендикулярно до осі ДНК. Перша впорядкована структура з постійною b1 зумовлює дифракційні рефлекси під кутом 1, які розташовані на паралельних шарових лініях. Відстань між цими лініями відповідає періоду b1 = 3,4 нм. Другий упорядкований елемент ДНК спричиняє інтенсивні меридіональні рефлекси, перший з яких збігається з 10-ю шаровою лінією й розташований під кутом 2. За наведеною вище формулою йому відповідає період b2 = 0,34 нм.
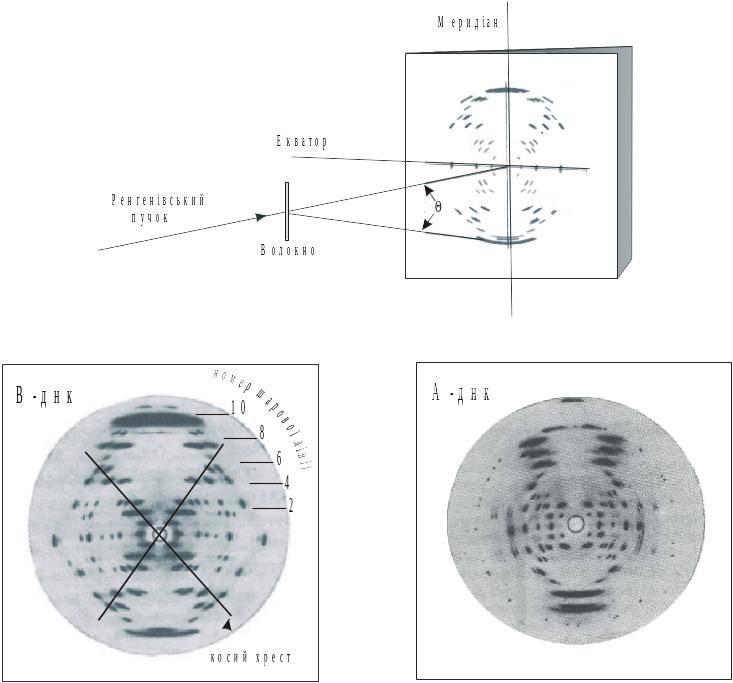
Рис. 5.10. Рентгенограми А- і В- форм ДНК
У запропонованій моделі пари основ складені у вигляді стовпчиків монет, відстань між якими становить 0,34 нм. При цьому сусідні пари повернуті на кут 360, унаслідок чого утворюється права подвійна спіраль, яка містить 10 пар основ на один оберт. Напрямок полінуклеотидних ланцюгів у спіралі ДНК антипаралельний. Головна позитивна характеристика моделі Уотсона–Кріка полягає в її адекватності реальності, у здатності пояснити однозначно механізм передачі спадкової інформації. Це реалізується завдяки принципу комплементарності нуклеотидних пар: аденін є комплементарним тиміну, і вони утворюють між собою два водневі зв’язки. Друга комплементарна пара гуанін – цитозин утворює три водневі зв’язки. Таким чином, кожна індивідуальна нитка ДНК містить повну інформацію про цілу молекулу. І це важливо не тільки для реалізації передачі спадкової інформації, але й для репарації (ремонту) пошкоджених під дією різних факторів ділянок ДНК, якщо вони належать тільки одному ланцюгу.