

Экзаменационный билет № 7
Основные типы жизненных циклов грибов, водорослей и высших растений.
Половые органы зигомицетов – зигоспорангии – образуются на гифах. После оплодотворения вырастает новая особь, в которой созревают споры. Споры высыпаются на землю и, прорастая, дают начало новым экземплярам гриба. Таким образом происходит чередование полового и бесполого размножения.
Споры другой группы грибов – аскомицетов (например, сморчков) – развиваются в особых сумках (асках), расположенных на нижней поверхности плодового тела гриба. Попадая в землю, споры прорастают в гифы; на них образуются половые органы – мужской (антеридий) и женский (аскогоний). Спермации из антеридия через соединительную трубку – трихогину – попадают в аскогоний, где происходит оплодотворение. Из оплодотворённой женской половой клетки вырастает гриб с новым плодовым телом, снова готовый к размножению.
У базидиомицетов споры созревают в гимениальных пластинках, расположенных в базидиях (на нижней поверхности шляпки плодового тела). Созревшие споры высыпаются и переносятся ветром, после чего попадают в почву и прорастают в гифы. Затем у большинства базидиомицетов сливаются клетки одной или нескольких гиф; на диплоидной грибнице прорастают новые плодовые тела.
Водоросли размножаются бесполым и половым путем.
Бесполое размножение осуществляется в двух формах. К вегетативному относится деление слоевища на части без каких-либо преобразований в клетках: деление клеток надвое у одноклеточных, распадение колоний, фрагментация у многоклеточных. Более специализированная форма бесполого размножения — спорообразование, при котором в специальных органах или внутри вегетативных клеток возникают неподвижные споры или подвижные, снабженные жгутиками зооспоры. Те и другие образуются в больших количествах и, прорастая, дают новую водоросль.
Половое размножение заключается в формировании специализированных половых клеток — мужских и женских гамет и их последующем слиянии с образованием зиготы. При половом размножении у разных видов водорослей гаметы бывают трех основных типов: половые клетки одинакового размера и формы (изогамия); неодинаковые по размерам и подвижности мужские и женские половые клетки (гетерогамия); крупная и неподвижная женская половая клетка и небольших размеров сперматозоид со жгутиками (оогамия). Мужские и женские гаметы могут развиваться на одном организме или на разных. После оплодотворения диплоидная зигота прорастает и образует новое слоевище. У гаплоидных видов первое деление зиготы редукционное. Все последующие клетки, возникающие в процессе роста, гаплоидны.
Примером чередования поколений у водорослей может служить бурая водоросль ламинария. Гаметы и споры красных водорослей лишены жгутиков и неподвижны. Оплодотворение осуществляется при пассивном переносе мужских половых клеток к женскому половому органу. У бурых водорослей встречаются все формы размножения.
Вегетативное размножение происходит при случайном отделении частей слоевища, только у некоторых видов для этого существуют специальные почки.
Споровое размножение осуществляется путем образования гаплоидных спор, развивающихся в гаплоидные растения полового поколения — гаметофиты. Половой процесс представлен тремя формами: изогамной, гетерогамной и оогамной. Зигота прорастает в диплоидное растение — спорофит. Спорофит образует подвижные зооспоры. У ламинарий гаметофит существует недолго, в то время как спорофит — многолетний.
Свой жизненный цикл — чередование спорофита и гаметофита — высшие растения унаследовали, вероятно, от своих водорослевых предков. Как известно, у водорослей наблюдаются самые различные взаимоотношения диплоидной и гаплоидной фаз жизненного цикла. Но у водорослевого предка высших растений диплоидная фаза была, возможно, развита больше, чем гаплоидная. В этой связи представляет большой интерес тот факт, что от древнейших и наиболее примитивных высших растений вымершей группы риниофитов в ископаемом состоянии достоверно сохранились только спорофиты. Скорее всего это можно объяснить тем, что гаметофиты у них были более нежные и менее развитые. Это ха рактерно также для огромного большинства ныне живущих растений. Исключение составляют только моховидные, у которых гаметофит преобладает над спорофитом.
Эволюция жизненного цикла высших растений шла в двух противоположных направлениях. У моховидных она была направлена в сторону возрастания самостоятельности гаметофита и его постепенного морфологического расчленения, потери самостоятельности спорофита и его морфологического упрощения. Самостоятельной, вполне автотрофной фазой жизненного цикла моховидных стал гаметофит, а спорофит низведен до степени органа гаметофита. У всех остальных высших растений самостоятельной фазой жизненного цикла стал спорофит, а гаметофит у них в течение эволюции постепенно уменьшался и упрощался. Максимальная редукция гаметофита связана с разделением полов. Миниатюризация и упрощение однополых гаметофитов происходили весьма ускоренными темпами. Гаметофиты очень быстро теряли хлорофилл, и развитие все чаще осуществлялось за счет питательных веществ, накопленных спорофитом.
Наибольшая редукция гаметофита наблюдается у семенных растений. Бросается в глаза, что как среди низших, так и среди высших растений все крупные и сложно устроенные организмы представляют собой спорофиты (ламинарии, фукусы, лепидодендроны, сигиллярии, каламиты, древовидные папоротники, голосеменные и древесные покрытосеменные).
Кроссинговер и его цитологический механизм. Доказательства физического обмена участками хромосом при кроссинговере (опыт К. Штерна, опыт Х.Крейтон и Б. Мак-Клинток).
Кроссинговер (crossing-over): обмен генетического материала между хромосомами, как результат "разрыва" и соединения хромосом; процесс обмена участками хромосом при перекресте хромосом (рис. 118 , Б4).
Во время пахитены (стадия толстых нитей), гомологичные хромосомы находятся в состоянии конъюгации длительный период: у дрозофилы - четверо суток, у человека больше двух недель. Все это время отдельные участки хромосом находятся в очень тесном соприкосновении. Если в таком участке произойдет разрыв цепочек ДНК одновременно в двух хроматидах, принадлежащих разным гомологам, то при восстановлении разрыва может получиться так, что ДНК одного гомолога окажется соединенной с ДНК другой, гомологичной хромосомы. Этот процесс носит -название кроссинговера (англ. crossing-over - перекрест).
Поскольку кроссинговер - взаимный обмен гомологичными участками хромосом между гомологичными (парными) хромосомами исходных гаплоидных наборов - особи имеют новые, различающиеся между собой генотипы. При этом достигается перекомбинация наследственных свойств родителей, что увеличивает изменчивость и дает более богатый материал для естественного отбора.
Гены перемешиваются благодаря слиянию гамет двух различных особей, однако генетические изменения осуществляются не только этим путем. Никакие два потомка одних и тех же родителей (если только это не идентичные близнецы) не будут абсолютно одинаковыми. Во время мейоза осуществляются два различных вида пересортировки генов.
Один вид пересортировки - это результат случайного распределения разных материнских и отцовских гомологов между дочерними клетками при первом делении мейоза, каждая гамета получает свою, отличную от других выборку материнских и отцовских хромосом. Из этого следует, что клетки любой особи могут в принципе образовать 2 в степени n генетически различающихся гамет, где n - гаплоидное число хромосом. Однако на самом деле число возможных гамет неизмеримо больше из-за кросинговера (перекреста) - процесса, происходящего во время длительной профазы первого деления мейоза , когда гомологичные хромосомы обмениваются участками. У человека в каждой паре гомологичных хромосом кроссинговер происходит в среднем в 2 - 3 точках .
При кроссинговере происходит разрыв двойной спирали ДНК в одной материнской и одной отцовской хроматиде, а затем получившиеся отрезки воссоединяются "наперекрест" (процесс генетической рекомбинации). Рекомбинация происходит в профазе первого деления мейоза, когда две сестринские хроматиды так тесно сближены друг с другом, что их невозможно увидеть в отдельности. Гораздо позже в этой растянутой профазе становятся ясно различимы две отдельные хроматиды каждой хромосомы. В это время видно, что они связаны своими центромерами и тесно сближены по всей длине. Два гомолога остаются связанными в тех точках, где произошел кроссинговер между отцовской и материнской хроматидами. В каждой такой точке, которую называютхиазмой , две из четырех хроматид перекрещиваются Таким образом, это морфологический результат произошедшего кроссинговера, который сам по себе недоступен для наблюдения.
Н а
этой стадии мейоза гомологи
в каждой паре (или бивалент) остаются
связанными друг с другом по меньшей
мере одной хиазмой. Во многих бивалентах
бывает большее число хиазм, так как
возможны множественные перекресты
между гомологами
а
этой стадии мейоза гомологи
в каждой паре (или бивалент) остаются
связанными друг с другом по меньшей
мере одной хиазмой. Во многих бивалентах
бывает большее число хиазм, так как
возможны множественные перекресты
между гомологами
Харриет Крейтон и Барбара МакКлинток решили эту проблему для кукурузы. Была исследована форма дигетерозиготная по генам (рис. 7.2): с (colorless) — бесцветный алейроновый слой эндосперма, рецессив; с! — окрашенный, доминант; wx (waxy) — эндосперм, а также пыльцевые зерна содержат амилопектин, вследствие чего окрашиваются иодом в красный цвет, рецессив; wx+ — эндосперм и пыльцевые зерна содержат амилозу, окрашиваемую иодом в синий цвет, доминант. Кроме того, исследованная форма на одном конце хромосомы несла блок (участок) гетерохроматина, а на другом — транслоцированный участок другой хромосомы. Второй гомолог этих морфологических отличий не имел. В соответствии со схемой рис. 7.2 в потомстве отбирали два реципрокных класса, рекомбинантных по с — wx. Растения, выращенные из семян этих рекомбинантов, исследовали цитологически и убедились в том, что у всех рекомбинантов по генетическим маркерам произошел физический обмен участками гомологичных хромосом (рис. 7.2).
Р ис.
7.2. Схема эксперимента X. Крейтон и Б.
МакКлинток, демонстрирующего соответствие
между рекомбинацией генов и физическими
обменами участками гомологичных
хромосом у кукурузы. Выделены
рекомбинантные классы потомства, в
которых исследовали структуру хромосом
ис.
7.2. Схема эксперимента X. Крейтон и Б.
МакКлинток, демонстрирующего соответствие
между рекомбинацией генов и физическими
обменами участками гомологичных
хромосом у кукурузы. Выделены
рекомбинантные классы потомства, в
которых исследовали структуру хромосом
Сходным образом решил ту же проблему К. Штерн для дрозофилы. Он исследовал расщепление в потомстве от скрещивания самок, дигетерозиготных по генам X- хромосомы: cr (carnation) глаза цвета гвоздики, рецессив; сг+ — нормальная темно-красная окраска глаз, доминант; В (Ваг) полосковидные глаза, доминант: В+ — круглые глаза, рецессив.
Эти же самки несли гетероморфные Х-хромосомы: одну сильно укороченную вследствие транслокации ее бесцентромерного участка на четвертую — микрохромосому, другую — Г-образную — вследствие транслокации на нее фрагмента Y-хромосомы. Таких самок скрещивали с самцами, несущими рецессивные аллели исследуемых генов (сг, В ) в Х-хромосоме обычной формы и имевшими обычную Y-хромосому (рис. 7.3). Результат кроссинговера у самок в таком скрещивании — рекомбинантные классы — изучали цитологически. Для этой цели брали только самок, поскольку самцы имеют субметацентрическую, т. е. разноплечую Y-хромосому, которую можно спутать с Г-образной Х-хромосомой.
Рис. 7.3. Схема эксперимента К. Штерна, демонстрирующего соответствие между рекомбинацией генов и физическими обменами участками Х-хромосом у дрозофилы.
В потомстве от скрещивания исследованы только самки (см. пояснение в тексте)

Благодаря этому, исследовав 374 самки, К. Штерн показал, что кроссинговер между генами сг и В сопровождался физическим обменом между Х-хромосомами.
Основные этапы процесса транскрипции. Посттранскрипционная модификация РНК. "Кэпирование", полиаденирование и сплайсинг мРНК у эукариот.
Транскрипцией называется перенос информации с двуцепочечной молекулы ДНК на одноцепочечные молекулы РНК. При этом матрицей для синтеза РНК служит только одна цепь ДНК, называемая смысловой цепью.
В транскрипции, так же, как и в других матричных процессах, различают три стадии: инициацию, элонгацию и термцнацию. Фермент (или ферменты), осуществляющий этот процесс, называют ДНК-зависимой РНК-полимеразой или просто РНК-полиме- разой, которая проводит следующую реакцию:

При этом полимеризация полирибонуклеотида (РНК) происходит в направлении от 5' к З'-концу растущей цепи.
Лучше всего изучена РНК-полимераза Е. coli. Большой вклад в расшифровку субъединичной структуры этого фермента внесла группа советских исследователей под руководством Р. Б. Хесина. РНК-полимераза Е. coli состоит из четырех субъединиц: двух одинаковых —α, которые объединены в молекуле так называемого минимального фермента еще с двумя различными субъединицами: (β и β!). Такой минимальный фермент (2αββ!)' с молекулярной массой 480 кДа осуществляет транскрипцию, но не способен «узнавать» на ДНК специфические стартовые позиции начала этого процесса. Для «узнавания» стартовых участков, или промоторов, к минимальному ферменту должна присоединяться еще одна субъединица — σ (сигма) - фактор.
Нуклеотидные
последовательности промоторов обычно
богаты парами AT. Их называют также
ТАТА-последовательностями или
последовательностями Прибнова. Примеры
некоторых из них представлены на рис.
15.8.
 После
инициации транскрипции σ-фактор
покидает минимальный фермент и дальнейшая
элонгация, т. е. наращивание РНК,
производится тетрамером РНК-полимеразы
(рис. 15.9).
После
инициации транскрипции σ-фактор
покидает минимальный фермент и дальнейшая
элонгация, т. е. наращивание РНК,
производится тетрамером РНК-полимеразы
(рис. 15.9).
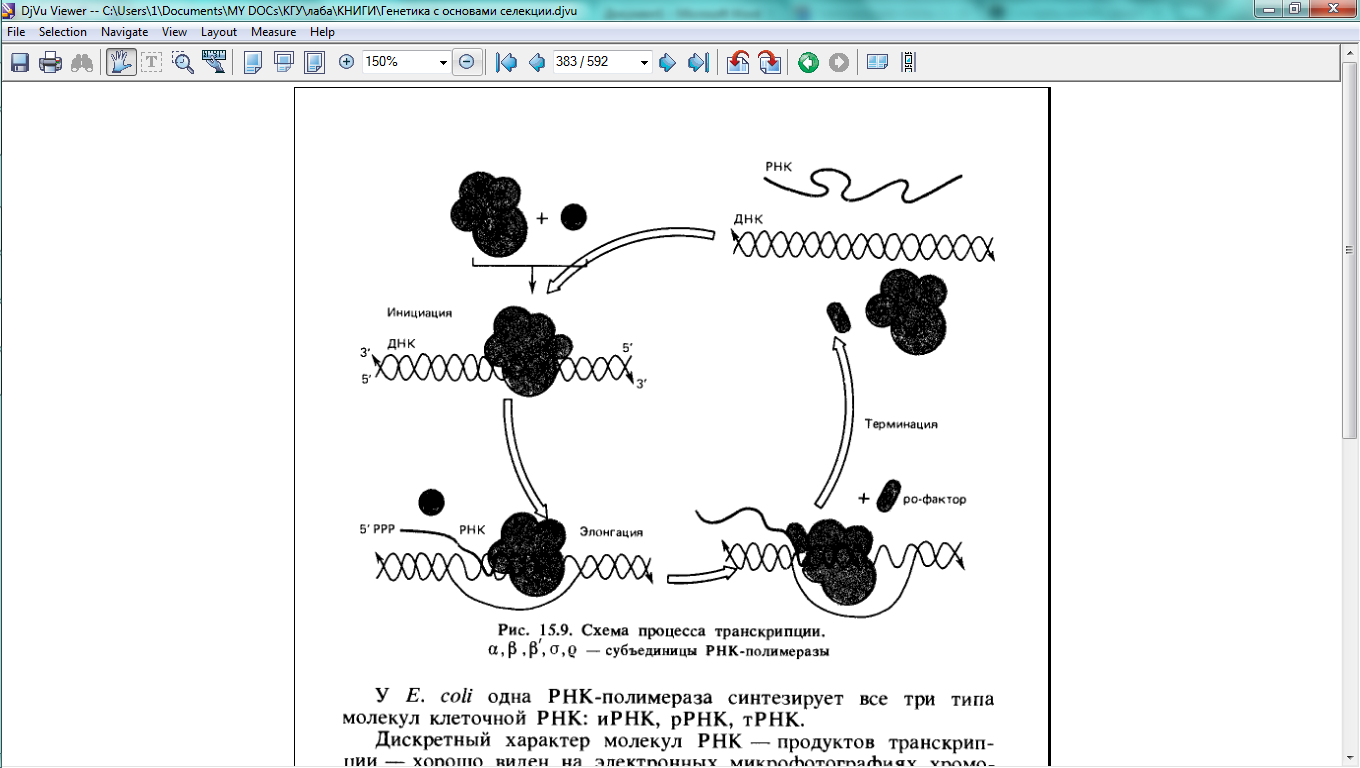
Скорость роста цепи РНК у Е. coli при 37°С составляет около 40—45 нуклеотидов в секунду. Завершается процесс транскрипции после присоединения другого белкового фактора— ρ (ро)-фактора, или фактора терминации (рис. 15.9). Терминация транскрипции происходит в специальных участках ДНК — терминаторах, содержащих инвертированные повторы.
У Е. coli одна РНК-полимераза синтезирует все три типа молекул клеточной РНК: иРНК, рРНК, тРНК.
Дискретный характер молекул РНК — продуктов транскрипции — хорошо виден на электронных микрофотографиях хромосом типа ламповых щеток (см. гл. 4).
У эукариот известны три типа РНК-полимераз: I — ответственная за синтез рРНК, II — ответственная за синтез иРНК, III — ответственная за синтез тРНК и низкомолекулярной рРНК — 5S РНК.
Поскольку РНК — продукт транскрипции — синтезируется на матрице ДНК, следует ожидать высокой степени корреляции нуклеотидного состава этих двух типов полимеров. Однако исследования, предпринятые в конце 50-х годов А. Н. Белозерским и А. С. Спириным, показали лишь небольшую корреляцию нуклеотидного состава РНК и ДНК из различных источников. Это указывало на то, что только небольшая фракция клеточной РНК в каждый данный момент отражает состав ее ДНК. Э. Волкин и Ф. Астрахан, изучая синтез РНК в бактериях Е. coli, зараженных фагом Т2, выявили в клетках новую фракцию РНК, состав которой подобен составу ДНК фага. Эта РНК была очень нестабильной. Так была открыта иРНК, переносящая информацию от ДНК к рибосомам, на которых синтезируется белок. Поскольку подавляющую часть клеточной РНК представляют стабильные рРНК и тРНК, общая корреляция состава ДНК и РНК оказывается незначительной. В дальнейшем у эукариот были открыты особые типы стабильной иРНК, например иРНК глобинов, присутствующая в безъядерных эритроцитах млекопитающих.
Посттранскрипционные модификации РНК особенно характерны для эукариот, у которых в силу мозаичной интрон-экзонной структуры их генов первичные транскрипты представлены гигантскими предшественниками, включающими в себя последовательности как экзонов, так и интронов. 5'-Конец предшественника мРНК чаще всего подвергается котранскрипционным модификациям, в результате которых к его 5'-концевому нуклеотиду особым образом присоединяется остаток гуанозина с образованием "шапочки" - кэпа. Эта котранскрипционная модификация создает условия для прохождения следующего этапа процессинга мРНК - сплайсинга, сопровождающегося вырезанием последовательностей интронов и объединением экзонов с образованием непрерывной кодирующей последовательности мРНК. Одновременно от 3'-конца путем эндонуклеазного расщепления отделяется избыточный фрагмент РНК, и к оставшейся части присоединяется поли(А)-последовательность. Эта совокупность реакций получила название полиаденилирования мРНК.
После таких котранскрипционных и посттранскрипционных модификаций пре-мРНК образовавшаяся зрелая, стабилизированная мРНК переносится из ядра в цитоплазму, часто к специфическому месту своей внутриклеточной локализации, где может быть депонирована или эффективно транслироваться рибосомами. Каждый из этапов посттранскрипционных модификаций может использоваться для регуляции уровня экспрессии соответствующих генов.