

25 Импеданс. Распределение основных компонентов импеданса в биологической ткани.
Электропроводность биологических систем – это качественная характеристика способности мембран, клеток, ткани и других. биологических объектов проводить электрический ток. В общем случае
![]()
где Z – импеданс ткани – ее полное cопротивление, равное сумме активного (R) и реактивного (XC+XL) сопротивлений: Z=R+XC+XL;
ХС = 1/ωС = 1/2πfC – емкостная составляющая импеданса;
ХL = ωL – индуктивная составляющая импеданса;
ω = 2πf – круговая частота подводимого переменного напряжения с частотой f.
Живой организм является проводником 2-го, так как направленный перенос зарядов в нём осуществляют ионы. От количества свободных ионов (переносчиков заряда) зависит электропроводность.
Для биологических объектов индуктивная составляющая L→0 и величина импеданса в выражении Z=R+XC+XL определяется суммой активной и емкостной составляющей Z=R+XC.
При параллельном соединении компонентов:
![]()
Для измерения импеданса применяют мостовые схемы (рис.1)
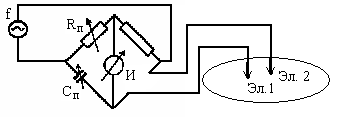
Рис. 1 – Применение мостовой схемы для регистрации импеданса тканей биологического объекта
В клетках основной катион – это К+, а кислоты (угольная, лимонная, уксусная, молочная) – основные анион.
Ионы биополимеров - белков, нуклеиновых кислот и их комплексов на величину электропроводности не влияют.
В межклеточной и межтканевой жидкости величина G определяется ионами Na+ и Cl-.
В тканях в результате переноса электронов и ионов от одних молекул к другим возникают окислительно-восстановительные потенциалы.
Величины электропроводности несут информация о функциональном состоянии тканей и органов, об изменениях наступающих под воздействием внешних факторов.
В медицине электропроводность характеризует патологические состояния: воспалительные процессы, изменения проницаемости сосудов (реография) и т.д.
На основе регистрации биоэлектрических потенциалов созданы методы электрокардиографии, электромиографии.
Изучение электрических процессов в нервной системе, например, потенциалов мозга занята нейроэлектрофизиология.
26 Преобразование информации в органе слуха. Слуховой анализатор: устройство и характеристики (роль стереоцилий – рецептора механических колебаний, чувствительность рецептора).
Слуховая система получает, перерабатывает и передает информацию. Слуховой анализатор — совокупность соматических, рецепторных и нервных структур, деятельность которых обеспечивает восприятие человеком и животными звуковых колебаний. С. а. состоит из наружного, среднего и внутреннего уха, слухового нерва, подкорковых релейных центров и корковых отделов.
Ухо является усилителем и преобразователем звуковых колебаний. Через барабанную перепонку, представляющую собой эластичную мембрану, и систему передаточных косточек — молоточек, наковальню и стремечко — звуковая волна доходит до внутреннего уха, вызывает колебательные движения в заполняющей его жидкости.
Внутреннее ухо, или улитка, представляет собой спиралеобразный ход, состоящий из двух с половиной витков. Заполняющая улитку жидкость — пери- и эндолимфа — практически несжимаема; поэтому при смещении стремечка вправо мембрана круглого окна прогибается влево, а возникающие колебания эндолимфы передаются волокнам расположенной вдоль улитки базилярной, или основной, мембраны и возбуждают специализированные механорецепторы — волосковые клетки. Улитка устроена так, что в одной ее части усиливаются звуковые волны высокой частоты, в другой – низкой частоты. Таким образом, улитка представляет собой подобие спектроанализатора: различные волосковые клетки отвечают на колебания различных частот.
Волосковые клетки улитки являются основными аппаратами слуховой рецепции. Реагируя на колебания эндолимфы, они превращают улавливаемые звуковые колебания в нервные импульсы, передающие акустическую информацию по волокнам слухового нерва.
Возбуждение, возникающее в волокнах слухового нерва, направляется к центральным отделам нервной системы. Первым центром обработки акустической информации являются расположенные на уровне варолиева моста ядра слухового нерва, после чего она поступает к т.н. верхним оливам. Здесь происходит объединение сигналов, поступающих от левой и правой улитки. Затем афферентные пути слухового нерва направляются к нижним буграм четверохолмия, которые представляют собой элементарный рефлекторный центр слуховой системы. Именно здесь осуществляется передача слуховых импульсов на двигательные пути, в результате чего возникают такие, напр., реакции, как двигательное настораживание или сокращение зрачка в ответ на внезапно возникающий звук.
Далее мощный пучок нервных волокон идет к внутренним коленчатым телам, от которых начинается последняя часть слухового нерва. Его волокна направляются к поперечной извилине височной области коры, или извилине Гешля, представляющей собой корковый конец. Вторичные отделы слуховой коры представляют собой сложный интегрирующий аппарат, который обеспечивает сложные формы анализа и синтеза звуковой информации, делая возможным восприятие сложномузыкальных и речевых звуков, поэтому поражение вторичных отделов слуховой коры не приводит к снижению остроты слуха и выпадению восприятия простых звуков, вызывает нарушение различения мелодий в одних случаях или сложно построенных звуков речи в других.
Преобразователем энергии механических колебаний в энергию электрического потенциала является волосковые клетки кортиева органа, расположенные на базилярной мембране.
Волосковые клетки, оканчиваются стереоцилиями. Стереоцилии - механочувствительные органеллы волосковых клеток длиной 3-мкм, расположенные на них в 3-4 ряда, связаны между собой внешним цитоскелетом. Кончики стереоцилий зафиксированы на неподвижной текториальной мембране. При звуковом воздействии, вызывающем вибрацию базилярной мембраны с расположенными на ней волосковыми клетками, стереоцилии изгибаются, в результате чего в них возникают рецепторные потенциалы – мембранный потенциал, модулированный затухающими колебаниями, частота которых совпадает с резонансной частотой, на которую настроена данная волосковая клетка. Этот потенциал синаптически возбуждает нервную клетку, по аксону которой электрические сигналы поступают в мозг.
Чувствительность волосковых клеток очень высока. Преобразующая система в ухе млекопитающего работает со звуками, в диапазоне интенсивностей, разнящихся по величине на семь порядков. Эта система позволяет слышать слабые звуки, вызывающие вибрацию базилярной мембраны с амплитудой, не превышающей доли диаметра водородного атома (порядка 10-3-10-4 нм).
27 Кровь, состав крови, роль системы кровообращения, ее функциональная и структурная организация (состав, функции элементов сердечно-сосудистой системы). Реологические свойства крови и режимы ее течения в различных элементах сердечно-сосудистой системы.
С ердечно-сосудистая система обеспечивает циркуляцию крови по замкнутой системе сосудов (рис 1.1). Постоянная циркуляция обеспечивает доставку во все клетки веществ, необходимых для их нормального функционирования и удаление продуктов жизнедеятельности.
Основная функция сердечно-сосудистой системы – обеспечение непрерывного движения крови по капиллярам, ответственным за обмен веществ между кровью и тканями. Для обеспечения процесса обмена веществ в капиллярах, сердечно сосудистая система имеет определенную функциональную и структурную организацию.
Состав и биофизические функции элементов сердечно-сосудистой системы
Сердце – насос, периодически выталкивающий кровь (в фазе систолы) в системы кровотока. Масса крови, выбрасываемой сердцем в аорту и далее в артерии в течение нескольких часов превышает массу человеческого тела. Следовательно, в сердце, играющем роль гидравлического насоса, многократно поступает одна и та же кровь. Крупные сосуды смягчают толчки – переводят пульсирующее давление в постоянное. Артерии запасают кровь во время систолы и выталкивают ее. В то же время эти крупные сосуды выполняют функцию проводников, подводя кровь к различным частям тела и органам.
Артериолы – резистивные сосуды. Легко изменяя свой просвет они регулируют гемодинамические показатели кровотока в капиллярах. Артериолы – «краны» сердечно-сосудистой системы. Это становится возможным благодаря эластичности крупных сосудов, которые по существу, являются согласующим звеном между сердцем и капиллярами. По венам кровь возвращается в сердце. Таким образом, в сердечно-сосудистой системе одновременно протекают разнородные процессы, взаимосвязанные друг с другом:
1. Поступление крови из левого желудочка в аорту и кровоток по сосудам.
2. Изменение давления крови и механических напряжений в стенках сосудов.
3. Изменение объема и формы элементов сердечно-сосудистой системы.
Гемодинамические показатели кровотока определяются биофизическими параметрами всей сердечно-сосудистой системы – как собственными характеристиками сердечной деятельности (например, ударным объемом крови), так и структурными характеристиками сосудов (их радиусом и эластичностью) и реологическими свойствами самой крови (прежде всего, ее вязкостью). Сердечно-сосудистая система – самосогласованная система со сложными взаимно-обратными связями.
Реологические свойства крови
Реология – наука о деформациях и текучести вещества. Гемореология – изучение биофизических особенностей крови как вязкой жидкости. Вязкость или внутреннее трение – свойство жидкости оказывать сопротивление перемещению одной ее части относительно другой. Вязкость жидкости обусловлена межмолекулярным взаимодействием, ограничивающим подвижность ее молекул. Наличие вязкости приводит к диссоциации (рассеянию и поглощению) энергии внешнего источника, вызывающего движение жидкости, и переходу ее в тепловую. Согласно основному закону вязкого течения (Ньютон 1867):
![]() (1)
(1)
где: F – сила внутреннего трения (вязкости), возникающей между слоями жидкости при сдвиге одного из них относительно другого;
η [Па/с] – коэффициент динамической вязкости жидкости, характеризующий сопротивление жидкости смещению ее слоев;
![]() [1/c]
– градиент скорости, показывающий, на
сколько изменяется скорость частицы
V
в потоке при переходе от слоя к слою
на единицу расстояния;
[1/c]
– градиент скорости, показывающий, на
сколько изменяется скорость частицы
V
в потоке при переходе от слоя к слою
на единицу расстояния;
S [м2]– площадь соприкасающихся слоев.
Сила внутреннего трения тормозит более быстрые слои и ускоряет более медленные.
Коэффициент кинематической вязкости
 (2) где
ρ
– плотность жидкости.
(2) где
ρ
– плотность жидкости.
По вязкости жидкости делятся на ньютоновские и неньютоновские. Ньютоновская – это такая жидкость, коэффициент вязкости, которой зависит только от ее природы и температуры. Для ньютоновской жидкости коэффициент вязкости прямо пропорционален градиенту скорости. Для нее справедлива формула Ньютона (1), в которой коэффициент вязкости η=const и не зависит от условий течения жидкости.
Неньютоновской называется жидкость, коэффициент вязкости которой зависит от природы вещества, температуры и условий течения жидкости (в частности, от градиента скорости). Для неньютоновской жидкости коэффициент вязкости не является константой вещества. Вязкость неньютоновской жидкости характеризуется условным коэффициентом вязкости, соответствующим определенным условиям течения жидкости, скорости, давлению и т.д. Сила вязкости зависит от градиента
скорости нелинейно:
 (3)
(3)
где n характеризует механические свойства при данных условиях течения. Пример неньютоновской жидкости – суспензии, в которых равномерно распределены твердые вещества. Если расстояния между частицами много больше диаметра частиц, среду можно рассматривать как однородную. Свойства такой среды зависят от η жидкости, но система в целом будет иметь вязкость η1 больше чем вязкость η. При малой концентрации частиц
![]() (4)
(4)
где к – геометрический фактор, зависящий от формы и размеров частиц
с – концентрация.
Кровь – неньютоновская жидкость. Она обладает внутренней структурой, представляя собой суспензию форменных элементов в растворе – плазме. Сама плазма – ньютоновская жидкость. 93% форменных элементов крови – эритроциты, поэтому кровь можно рассматривать как суспензию эритроцитов в физиологическом растворе.
Характерным свойством крови является образование агрегатов: эритроциты склеиваются друг с другом, образуя агрегаты, которые называются «монетными столбиками». В крупных и мелких сосудах условия агрегатирования различны. Это связано с соотношением размеров сосудов (dсос), агрегата (dагр) и эритроцита (dэр), для которых dэр= 8 мкм, dагр =10dэ.Здесь возможны различные варианты:
В
крупных сосудах
(аорты, артерия) dсос>dагр;
dсос>>dэр
и градиент
![]() небольшой. Эритроциты
собираются в виде монетных столбиков.
В этом случае вязкость крови η=0,005 [Па
с].
небольшой. Эритроциты
собираются в виде монетных столбиков.
В этом случае вязкость крови η=0,005 [Па
с].
В мелкие сосудах (мелкие артерии). dсос=dагр; dсос=(5-20) dэр градиент значительно увеличивается, и агрегаты распадаются на отдельные эритроциты, уменьшая тем самым вязкость системы. Кровь представляет собой суспензию форменных элементов. Вязкость крови составляет примерно 2/3 вязкости крови в крупных сосудах.
Микрососуды. Эритроциты, не разрушаясь, легко проходят капилляр диаметром до 3 мкм, в котором они не деформируются и становятся похожими на купол. Поверхность соприкосновения эритроцита с капилляром увеличивается по сравнению с недеформированным эритроцитом, и в результате, процесс обмена веществ ускоряется.Если предположить, что в случаях 1 и 2 эритроциты не деформируются, то для качественного описания изменения вязкости применима формула , в которой можно учесть различие геометрического фактора для системы из агрегатов и системы из отдельных эритроцитов, обуславливающее различие вязкости в крови крупных и мелких сосудов. Вязкость возрастает при снижении температуры на 10% при уменьшении температуры с 370С до 170С
Режимы течения крови
С увеличением скорости движения ламинарное движение кровотока переходит в турбулентное, при котором происходит интенсивное перемешивание между слоями жидкости. В потоке возникают многочисленные вихри различных размеров. Частицы совершают хаотическое движение по сложным траекториям. Для турбулентного течения характерны нерегулярные изменения скорости со временем в каждой точке потока.
Скорость турбулентного течения в трубах отличается от параболического профиля ламинарного течения более быстрым возрастанием скорости у стенок и меньшей кривизной в центральной части течения. Профиль скоростей при этом описывается логарифмическим законом.
Режим течения жидкости характеризуется числом Рейнольдса (Re). Для круглой трубы
![]() ,
(6)
,
(6)
где V – средняя по поперечному течению скорость течения,
R – радиус трубы.
При Re <2300 (критическое значение) – движение ламинарное.
При Re>Rкр – турбулентное.
Турбулентность кровотока возникает в местах сужения сосудов, например, при тромбозе, при прохождении крови из желудочка в аорту. Турбулентность возникает в местах разветвления сосудов, а также в самих артериях при возрастании скорости движения в крови, например, при мышечной нагрузке или при образовании тромба. Шум, возникающий при турбулентности, можно регистрировать и по нему судить о заболеваниях. Например, при поражении клапанов сердца, возникают сердечные шумы, вызванные турбулентным движением крови.
28 Мышечная ткань, ее функции. Роль мышечной активности в жизнеобеспечении высокоорганизованных живых организмов. Типы мышечных клеток, их структуры и функции, принадлежность к различным органам. Устройство и принцип работы сократительного аппарата. Режимы мышечного сокращения и их характеристики.
Мышечная активность – одно из общих свойств высокоорганизованных живых организмов. Вся жизнедеятельность человека связана с его мышечной активностью. Независимо от назначения, особенностей строения и способов регуляции принцип работы различных мышц организма одинаков.
Мышечная клетка отличается от других клеток возбудимой ткани – нейронов – специфическим свойством: способностью сокращаться – создавать механическое напряжение и укорачиваться. Кроме того, мышца генерирует тепло. В процессе жизнедеятельности мышечная активность обеспечивает работу отдельных органов опорно-двигательного аппарата, желудочно-кишечного тракта, сосудистую активность, сократительную способность сердца и легких.
Мышечная ткань представляет собой совокупность мышечных клеток, внеклеточного вещества (коллаген, элластин) и густой сети нервных волокон и кровеносных сосудов.
Различают 3 типа мышечных клеток:
1. Скелетная поперечнополосатая мышца сильно и быстро сокращаясь, обеспечивает движение сустава. Каждое ее волокно представляет собой огромную многоядерную клетку.
В основе управления клеток лежит нейрогенная регуляция, осуществляемая соматическим отделом нервной системы. Управление осуществляется при участии сознания.
Характер активности – как уже было отмечено ранее имеет высокое быстродействие; относительно короткий период рефрактерности; мощные сокращения; средняя работоспособность при статических и динамических нагрузках; относительно быстрое утомление.
Дина клеток поперечнополосатых мышц составляет ~1÷40 мм, диаметр - ~10÷60 мкм. В цитоплазме клеток расположены метоходрии (рядами по периферии волокон и между ними), сложный саркоплазматический ретикулум, саркотубулярная система. Клетки многоядерные. Ядра расположены у внутренней поверхности сложноорганизованной мембраны волокна - сарколеммы. Вставочные диски отсутствуют, однако хорошо видны нити актина и миозина.
2. Гладкие мышцы – это мышцы желудочно-кишечного тракта, мочевого пузыря и кровеносных сосудов. Они состоят из тонких одноядерных клеток, не имеющих исчерченности.
В основе управления клеток лежит миогенная регуляция. При управлении сознание не участвует.
Характер активности – низкое быстродействие, автоматическая ритмичность сокращений и расслаблений. Дина клеток гладких мышц составляет ~0,015÷0,5 мм, диаметр 2÷10 мкм. В цитоплазме клеток расположены хорошо выраженные митохондрии, очень простой эндоплазматический ретикулум. Клетки имеют одно овальное ядро, расположенное в центре клетки. Вставочные диски отсутствуют, нити актина и миозина плохо различимы.
3. Сердечная мышца занимает промежуточное положение между поперечнополосатыми и гладкими мышцами.
В основе управления лежит миогенная регуляция, нейрогенная решуляция с участием вегетативного отдела нервной системы, в процессе управления сознание практически не участвует.
Характер активности – высокое быстродействие, автоматическая ритмичность сокращений и расслаблений, относительно продолжительный рефрактерный период, высокая работоспособность и относительно медленное утомление при ритмических динамических нагрузках, низкая работоспособность и относительно быстрое утомление при статических нагрузках. Длина клеток ~0,08 мм или менее, диаметр ~12÷15 мкм.
Цитоплазма содержит многочисленные крупные митохондрии, сосредоточенные в саркоплазме у полюсов ядра, более простой саркоплазматический ретикулум, саркотубулярная система. Клетка имеет одно или два ядра, расположенных в центре клетки.
Клетки характеризуются наличием вставочных дисков и хорошей различимостью нитей актина и миозина.
Клетки поперечнополосатой и гладкомышечной ткани представлены на рисунке 1.
Рисунок 1 - Клетки поперечнополосатой и гладкомышечной ткани
Все мышцы имеют близкие механические свойства, одинаковый механизм активации и близкий химический состав.
Мышечное волокно – сильно вытянутая клетка диаметром 20-80 мкм, окруженная мембранной толщиной 10 нм. Длина волокон зависит от вида мышцы и составляет от сотен мкм до нескольких сантиметров. Кроме стандартного набора органелл (ядро, ядрышко, митохондрии, аппарат Гольджи и т.д.), имеется сократительный аппарат, состоящий из 1000-2000 параллельно идущих миофибрилл диаметром 1-2 мкм, а также саркоплазматический ретикулум (СР) и система поперечных трубок – Т-система (рис. 2.1).
В миофибриллах различают А-зону (темные полосы, обладающие свойством анизотропии) и изотропную I-зону – светлые полосы. В области I-зоны проходит поперечная темная узкая полоса – Z-диск.
Промежуток между двумя дисками, назывемый саркомером, является элементарной сократительной единицей мышечной клетки.
Саркомер – упорядоченная система толстых и тонких продольно расположенных нитей. Толстая нить, построенная из белка миозина, имеет диаметр 12 нм и длину 1,5 мкм.
Диаметр тонкой нити – 8 нм, длина – 1 мкм. Она построена из белка актина (2 мономера актина толщиной по 5 нм, закрученных один вокруг другого) и напоминает нитку бус из 14 бусин в витке. Одним концом она прикреплена к
Z-диску (рис.2).
В цепях актина регулярно, примерно через каждые 40 нм, встроены молекулы тропонина, а сама цепь охватывает нить тропомиозина. При сокращении мышцы тонкие нити актина вдвигаются между толстыми нитями миозина.
Рис.2 – Микроструктура саркомера Рис. 3 - Процесс активации мостика и генерации усилия в саркомере
Нити скользят, но свою длину не изменяют. Этот процесс обусловлен взаимодействием особых выступов миозина – поперечных мостиков с активными центрами, расположенными на актине. Мостики отходят от толстой нити на расстоянии 14,5 нм друг от друга. В расслабленном состоянии миофибрилл молекулы тропомиозина блокируют прикрепление поперечных мостиков к актиновым цепям (рис. 3а). Ионы Са+ активируют мостик и открывают участок его крепления к актину (рис.3б). В результате мостик миозина прикрепляется к актиновым нитям. При этом расщепляются молекулы АТФ и изменяется конформация мостика так, что его головки поворачиваются внутрь саркомера (рис. 3в). Это приводит к скольжению актиновой нити относительно толстой нити миозина в сторону центра саркомера и к укорочению мышцы, в результате которого возникает усилие.
После окончания активации мостики размыкаются, и саркомер возвращается в исходное состояние. При сокращении длины саркомера его объем не изменяется, происходит утолщение.
Каждый цикл замыкания-размыкания мостика сопровождается расщеплением 1 молекулы АТФ. Таким образом, актин-миозиновый комплекс является преобразователем энергии АТФ. Структура саркомера и последовательность происходящих в нем процессов называется “моделью скользящих нитей”. Основные положения модели скользящих нитей:
1. длины нитей миозина и актина в процессе сокращения мышцы не изменяются;
2. изменение длины саркомера при сокращении является результатом продольного смещения нитей актина и миозина относительного друг друга;
3. поперечные мостики, отходящие от миозина, присоединяются к комплементарным центрам актина;
4. мостики прикрепляются к актину не одновременно;
5. замкнувшиеся мостики подвергаются структурному изменению, при котором они развивают усилия, после чего происходит их размыкание;
6. сокращение и расслабление мышцы заключается в нарастании и последующем уменьшении числа мостиков, совершающих цикл замыкание-размыкание;
7. каждый цикл связан с гидролизом 1 молекулы АТФ;
8. акты замыкания-размыкания мостиков происходят независимо друг от друга.
На рис. 4.a представлена зависимость максимального значения развиваемой силы от степени перекрытия актиновых и миозиновых нитей. Если саркомер идеально растянут до длины 3,65 мкм, мостики не контактируют с актиновыми нитями и при стимуляции такого элемента усилие не возникает (стрелка 1 на рис. 4.а и фрагмент 1 на рис. 4.б). Если саркомер находится в начальном рабочем состоянии (размер саркомера 2,2 мкм), то при стимуляции он разовьет максимальную силу (стрелка 2 на рис. 4.а и фрагмент 2 на рис. 4.б). При дальнейшем уменьшении начальной длины саркомера, усилие, развиваемое им, спадает до 0 (стрелка и фрагмент 5 на рис. 4.а и 4.б).
Рис. 4.а- Зависимость максимального значения силы Р, Рис. 4.б - Зависимость максимального значения силы Р,
развиваемой при изометрическом сокращении: развиваемой при изометрическом сокращении от перекрытия
а. – от начальной длины саркомера; актиновых и миозиновых нитей
Мышцу можно представить как сплошную среду, состоящую из большого числа частиц, взаимодействующих между собой без соударений и находящихся в поле внешних сил. Мышцы обладают одновременно свойством упругости и вязкости, т.е. является вязкоупругой средой. Как вязкоупругое тело мышца характеризуется величинами деформации, напряженности, упругости, вязкости, температуры и энергии.
Упругость – свойство тел изменять размеры и форму под воздействием внешних сил и самопроизвольно восстанавливать их при снятии внешней нагрузки. Упругость тела обусловлена силами взаимодействия его атомов и молекул. При снятии внешнего воздействия тело возвращается в исходное состояние.
Вязкость – внутреннее трение среды. Для мышцы вязкость обусловлена скольжением нитей актина относительно миозина.
Деформация
– относительное
изменение начальной длины тела (мышцы)
на величину Δl:
![]()
Механическое
напряжение σ – мера
внутренних сил, возникающих при
деформации мышцы
![]() где F
– приложенная сила, S
– площадь сечения.
где F
– приложенная сила, S
– площадь сечения.
Упругая
деформация –
деформация, возникающая при приложении
усилия и исчезающая при его снятии.
Упругая деформация подчиняется закону
Гука:![]() где
ε – модуль Юнга. При малых растяжениях
ε =Сonst.
Для различных материалов модуль Юнга
равен: эластин – 6.105
Па;
коллаген – 109Па;
кость – 1010.
где
ε – модуль Юнга. При малых растяжениях
ε =Сonst.
Для различных материалов модуль Юнга
равен: эластин – 6.105
Па;
коллаген – 109Па;
кость – 1010.
Эластин – упругий белок позвоночных, находится в основном в стенках артерий.
Коллаген – волокнистый белок. В мышцах 20% от всех белков – коллаген. Он находится также в сухожилиях, хрящах, костях.
Для вязкой среды напряжение определяется скоростью деформации: σв = η dε/dt , где η – коэффициент вязкости среды.
В
общем случае для вязкоупругой среды
σ=σупр
+ σв=εЕ+
η dε/dt
откуда, решая
дифференциальное уравнение при t
= 0 и ε
= 0 получим:
ε(t)=
σ/Е(1-е-![]() ),
где величина τ = η/
Е называется временем
запаздывания.
),
где величина τ = η/
Е называется временем
запаздывания.
Для характеристики мышцы используется два искусственных режима
В
изометрическом
режиме длина мышцы остается постоянной
(![]() =Сonst),
регистрируется развиваемое мышцей
усилие F(
).
С этой целью с помощью фиксатора
предварительно устанавливают длину
мышцы (рис. 5а), после чего на электроды
подается стимул. Величину развившегося
усилия F
регистрируют с помощью датчика Д F..
=Сonst),
регистрируется развиваемое мышцей
усилие F(
).
С этой целью с помощью фиксатора
предварительно устанавливают длину
мышцы (рис. 5а), после чего на электроды
подается стимул. Величину развившегося
усилия F
регистрируют с помощью датчика Д F..
В изотоническом режиме рисунок 5.б усилие F=const; исследуется изменение длины мышцы во времени ∆ = f(t).


Рисунок 5 -Схемы опытных установок для реализации в эксперименте Рис. 6. Временная зависимость одиночного
а – изометрического, б – изотонического режимов. ДF – датчик силы; сокращения при: А-изометрическом и Б –
ДL – датчик изменения длины; М – мышца; Р – нагрузка; Эл – электрод изотоническом режимах сокращения мышцы
На рис. 6 показана зависимость F( ) в изометрическом режиме для двух мышц различной длины 1 и 2. Максимальная сила, развиваемая мышцей, зависит от начальной длины и области перекрытия актиновых и миозиновых нитей, в которой могут замыкаться мостики. При начальной длине саркомера о=2,2 мкм в сокращении участвуют все мостики (рис. 4б). Поэтому максимальное усилие F0, которое может развить мышца, возникает при таком предварительном растяжении, при котором длина саркомера равна 2,2 мкм. Как видно на рис.6.а, усилие F01 >F02 поскольку длина мышцы 1> 2 и, следовательно, в ее сокращении участвует большее число содержащихся в ней саркомеров.
При изотоническом режиме к незакрепленному концу мышцы привешивается груз Р. После этого подается стимул и регистрируется изменение длины мышцы во времени ∆ (t) (рис. 6б). Из рис. 6б следует, что с увеличением груза Р снижается как укорочение мышцы, так и продолжительность ее сокращения. При некотором значении Р=Р0 сокращение мышцы ∆ (или относительное изменение ее длины ε) равно нулю. Р0 – максимальная сила изотонического сокращения для данной мышцы. Скорость сокращения мышцы также падает с увеличением нагрузки.