

21 Роль мышечной активности в жизнеобеспечении высокоорганизованных живых организмов. Типы мышечных клеток. Структурное и функциональное различие клеток в различных органах.
Мышечная активность – одно из общих свойств высокоорганизованных живых организмов. Вся жизнедеятельность человека связана с его мышечной активностью. Независимо от назначения, особенностей строения и способов регуляции принцип работы различных мышц организма одинаков.
Мышечная клетка отличается от других клеток возбудимой ткани – нейронов – специфическим свойством: способностью сокращаться – создавать механическое напряжение и укорачиваться. Кроме того, мышца генерирует тепло. В процессе жизнедеятельности мышечная активность обеспечивает работу отдельных органов опорно-двигательного аппарата, желудочно-кишечного тракта, сосудистую активность, сократительную способность сердца и легких.
Мышечная ткань представляет собой совокупность мышечных клеток, внеклеточного вещества (коллаген, элластин) и густой сети нервных волокон и кровеносных сосудов.
Различают 3 типа мышечных клеток:
1. Скелетная поперечнополосатая мышца сильно и быстро сокращаясь, обеспечивает движение сустава. Каждое ее волокно представляет собой огромную многоядерную клетку.
В основе управления клеток лежит нейрогенная регуляция, осуществляемая соматическим отделом нервной системы. Управление осуществляется при участии сознания.
Характер активности – как уже было отмечено ранее имеет высокое быстродействие; относительно короткий период рефрактерности; мощные сокращения; средняя работоспособность при статических и динамических нагрузках; относительно быстрое утомление.
Дина клеток поперечнополосатых мышц составляет ~1÷40 мм, диаметр - ~10÷60 мкм. В цитоплазме клеток расположены метоходрии (рядами по периферии волокон и между ними), сложный саркоплазматический ретикулум, саркотубулярная система. Клетки многоядерные. Ядра расположены у внутренней поверхности сложноорганизованной мембраны волокна - сарколеммы. Вставочные диски отсутствуют, однако хорошо видны нити актина и миозина.
2. Гладкие мышцы – это мышцы желудочно-кишечного тракта, мочевого пузыря и кровеносных сосудов. Они состоят из тонких одноядерных клеток, не имеющих исчерченности.
В основе управления клеток лежит миогенная регуляция. При управлении сознание не участвует.
Характер активности – низкое быстродействие, автоматическая ритмичность сокращений и расслаблений. Дина клеток гладких мышц составляет ~0,015÷0,5 мм, диаметр 2÷10 мкм. В цитоплазме клеток расположены хорошо выраженные митохондрии, очень простой эндоплазматический ретикулум. Клетки имеют одно овальное ядро, расположенное в центре клетки. Вставочные диски отсутствуют, нити актина и миозина плохо различимы.
3. Сердечная мышца занимает промежуточное положение между поперечнополосатыми и гладкими мышцами.
В основе управления лежит миогенная регуляция, нейрогенная решуляция с участием вегетативного отдела нервной системы, в процессе управления сознание практически не участвует.
Характер активности – высокое быстродействие, автоматическая ритмичность сокращений и расслаблений, относительно продолжительный рефрактерный период, высокая работоспособность и относительно медленное утомление при ритмических динамических нагрузках, низкая работоспособность и относительно быстрое утомление при статических нагрузках. Длина клеток ~0,08 мм или менее, диаметр ~12÷15 мкм.
Цитоплазма содержит многочисленные крупные митохондрии, сосредоточенные в саркоплазме у полюсов ядра, более простой саркоплазматический ретикулум, саркотубулярная система. Клетка имеет одно или два ядра, расположенных в центре клетки.
Клетки характеризуются наличием вставочных дисков и хорошей различимостью нитей актина и миозина.
Клетки поперечнополосатой и гладкомышечной ткани представлены на рисунке 1.
Рисунок 1 - Клетки поперечнополосатой и гладкомышечной ткани
22 Устройство саркомера. Специфическое свойство и принцип работы сократительного аппарата.
Саркомер – упорядоченная система толстых и тонких продольно расположенных нитей. Толстая нить, построенная из белка миозина, имеет диаметр 12 нм и длину 1,5 мкм.
Диаметр тонкой нити – 8 нм, длина – 1 мкм. Она построена из белка актина (2 мономера актина толщиной по 5 нм, закрученных один вокруг другого) и напоминает нитку бус из 14 бусин в витке. Одним концом она прикреплена к
Z-диску (рис.1).
В цепях актина регулярно, примерно через каждые 40 нм, встроены молекулы тропонина, а сама цепь охватывает нить тропомиозина. При сокращении мышцы тонкие нити актина вдвигаются между толстыми нитями миозина.
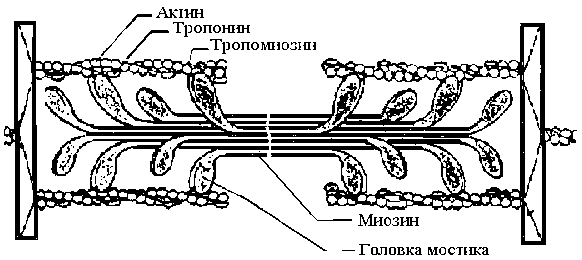
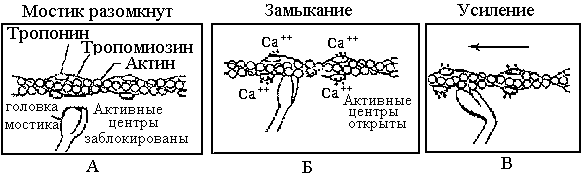
Рис.1 – Микроструктура саркомера Рис. 2 - Процесс активации мостика и генерации усилия в саркомере
Нити скользят, но свою длину не изменяют. Этот процесс обусловлен взаимодействием особых выступов миозина – поперечных мостиков с активными центрами, расположенными на актине. Мостики отходят от толстой нити на расстоянии 14,5 нм друг от друга. В расслабленном состоянии миофибрилл молекулы тропомиозина блокируют прикрепление поперечных мостиков к актиновым цепям (рис. 2.3а). Ионы Са+ активируют мостик и открывают участок его крепления к актину (рис.2.3б). В результате мостик миозина прикрепляется к актиновым нитям. При этом расщепляются молекулы АТФ и изменяется конформация мостика так, что его головки поворачиваются внутрь саркомера (рис. 2.3в). Это приводит к скольжению актиновой нити относительно толстой нити миозина в сторону центра саркомера и к укорочению мышцы, в результате которого возникает усилие.
После окончания активации мостики размыкаются, и саркомер возвращается в исходное состояние. При сокращении длины саркомера его объем не изменяется, происходит утолщение.
Каждый цикл замыкания-размыкания мостика сопровождается расщеплением 1 молекулы АТФ. Таким образом, актин-миозиновый комплекс является преобразователем энергии АТФ. Структура саркомера и последовательность происходящих в нем процессов называется “моделью скользящих нитей”. Основные положения модели скользящих нитей:
1. длины нитей миозина и актина в процессе сокращения мышцы не изменяются;
2. изменение длины саркомера при сокращении является результатом продольного смещения нитей актина и миозина относительного друг друга;
3. поперечные мостики, отходящие от миозина, присоединяются к комплементарным центрам актина;
4. мостики прикрепляются к актину не одновременно;
5. замкнувшиеся мостики подвергаются структурному изменению, при котором они развивают усилия, после чего происходит их размыкание;
6. сокращение и расслабление мышцы заключается в нарастании и последующем уменьшении числа мостиков, совершающих цикл замыкание-размыка- ние;
7. каждый цикл связан с гидролизом 1 молекулы АТФ;
8. акты замыкания-размыкания мостиков происходят независимо друг от друга.
На рис. 3 представлена зависимость максимального значения развиваемой силы от степени перекрытия актиновых и миозиновых нитей. Если саркомер идеально растянут до длины 3,65 мкм, мостики не контактируют с актиновыми нитями и при стимуляции такого элемента усилие не возникает (стрелка 1 на рис. 3.а и фрагмент 1 на рис. 3.б). Если саркомер находится в начальном рабочем состоянии (размер саркомера 2,2 мкм), то при стимуляции он разовьет максимальную силу (стрелка 2 на рис. 3.а и фрагмент 2 на рис. 3.б). При дальнейшем уменьшении начальной длины саркомера, усилие, развиваемое им, спадает до 0 (стрелка и фрагмент 5 на рис. 3.а и 3.б).
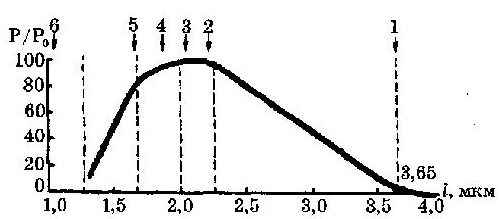
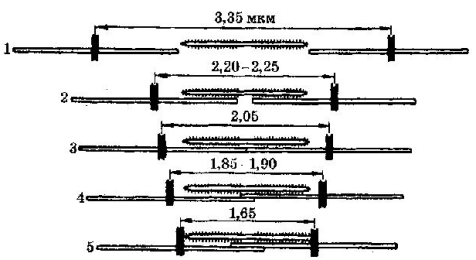
Рис. 3.а- Зависимость максимального значения силы Р, Рис. 3.б - Зависимость максимального значения силы Р,
развиваемой при изометрическом сокращении: развиваемой при изометрическом сокращении от перекрытия
а. – от начальной длины саркомера; актиновых и миозиновых нитей