

25. Основные этапы мышечного сокращения. Скольжение толстых и тонких нитей, мостиковая гипотеза генерации силы. Кинетическая теория мышечного сокращения в.И. Дещеревского.
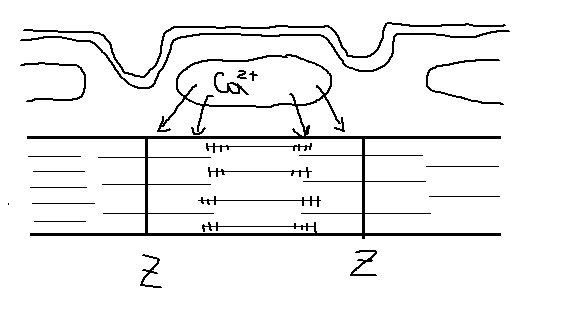
Этапы:
1. Деполяризация мембранного мышечного волокна ->
2. Высвобождение Ca2+ из саркоплазматического ретикулума (С=1мМ) -> в мышечную клетку(С0=10-7М).
3. Тропонин обладает высоким сродством к ионам Ca2+ -> конформация. Тропомиозиновая нить сдвигается и открывается активные центры на F-актине.
4. Взаимодействие миозиновых мостиков с активными центрами (актиновыми нитями).
5. Продвижение актиновых нитей относительно толстых миозиновых (на 10 нм за 1 состыковку).
6. Отсоединение центров на актиновых нитях и миозиновых мостиках. Благодаря АТФазной активности молекул миозина. Она способна вызывать гидрому молекулы АТФ и высвобождение энергии, за счет чего происходит разрыв связи актинового мостика с миозиновой нитью.
7. Если не повторять цикл (расслабленные мышцы), активируется работа Ca-насоса. Меньше всего ионов Ca2+ покидают клетку за счет сарколеммы. Большая часть – закачиваются в саркоплазматический ретикулум.
Миофибриллы (сократительный аппарат) состоят из тонких актиновых и толстых миозиновых нитей.
 ,
l~1,5мкм
,
l~1,5мкм
в состоянии миозиновых нитей – белок миозин
 ,
l~1мкм
,
l~1мкм
В составе актиновых нитей:
- актин
- тропомиозин
- тропонин: 3 субъединицы, 2 молекулы на 1 шаге
Функция тропомиозина – на нем активируются центры связывания с мостиками толстых нитей.
Миозиновая нить (одна из самых больших структур в природе)
 миозиновый
мостик обеспечивает скольжение нитей
друг относительно друга.
миозиновый
мостик обеспечивает скольжение нитей
друг относительно друга.
Кинетическая теория мышечного сокращения.
Основные положения:
Элементарная сила (усилие) развиваемая отдельным мостиком, постоянна и положительна. Направлена к центру саркомера на всем протяжении тянущего состояния.
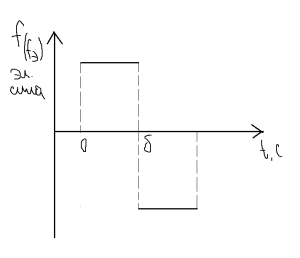
В тормозящем замкнутом состоянии: развиваемая мостиком в тормозящем состоянии сила имеет тот же значение, но она направлена противоположено (отрицательная).
Каждый поперечный мостик в процессе сокращения проходит через 3 состояния:
- свободно разомкнутое (среднее положение),
- тянущее замкнутое состояние, во время которого возникает тянущая сила f (+, в центр С)
- замкнутое тормозящее состояние, при котором развивается тормозящая сила f, которая препятствует скольжению нитей.
γ – количество свободных мостиков,
n – тянущее замкнутое состояние,
m – количество мостиков в замкнутом состоянии.
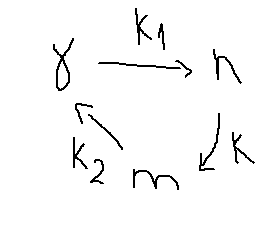
3) Переходы между состояниями для любого мостика не зависит от состояния других мостиков. Описываются реакциями первого порядка с константами скорости k1, k и k2. Эти константы могут зависеть от скорости скольжения нитей.
4) Константа скорости любого превращения первого порядка есть величина обратная времени, в течение которого данная структура существует до момента превращения. Время жизни мостика в тянущем состоянии: перемещение делить на скорость перемещения:


 - путь, который
проходит мостик в тянущем состоянии.
- путь, который
проходит мостик в тянущем состоянии.
U – скорость движения нитей друг относительно друга.
 ,
k1
не зависит от скорости скольжения U
,
k1
не зависит от скорости скольжения U
k2 и f не зависят от скорости скольжения нитей,
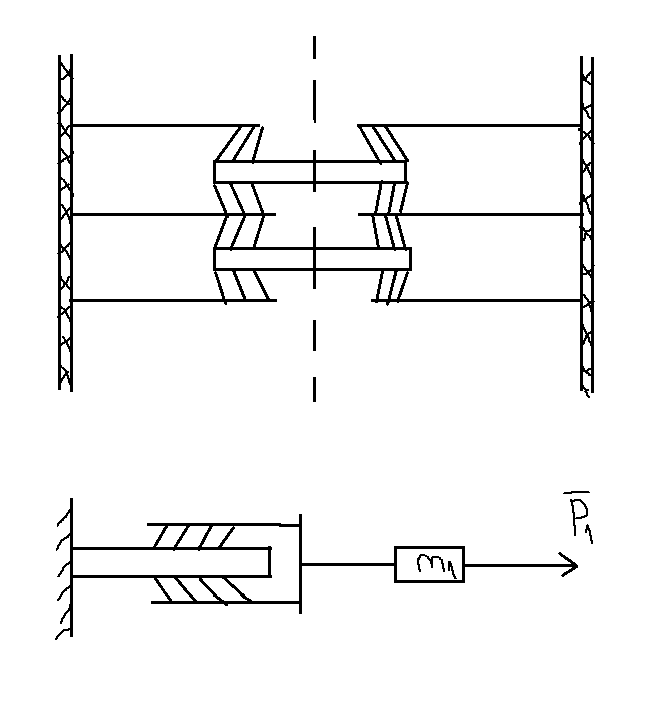


m1 – условная часть массы груза M, приходящаяся на 1 миозиновую нить в полу-саркомере.
f
– сила,
развиваемая одним мостиком,

Р1 – часть внешней нагрузки Р, приходящаяся на эту саркомерную нить.
а, V1 – ускорение и скорость сокращающегося полусаркомера.
(n-m) – разность между количеством тянущих и тормозящих мостиков.
(n-m)f – тянущая сила одной миозиновой нити в полусаркомере.
Р0 – максимальное усилие, развиваемое в мышце при изолированном сокращении.
 ,
[a]=[P]=H
,
[a]=[P]=H
 ,
[b]=[V]=м/с
,
[b]=[V]=м/с
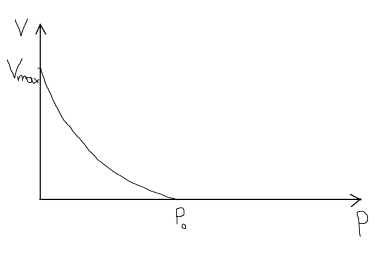

площадь сечения мышцы:
 (штук
мостиков)
(штук
мостиков)

За один поворот мостиков:

Активная мышца растягиваться не может.