

22. Поперечно-полосатая мускулатура, структура сократительного аппарата, молекулярная организация.
Структура Сократительного аппарата:
Миофибриллы (белковая структура, нити толстые и тонкие)
Миофибриллы d≈1000-2000 мф-л
Сократительная единица- сакромер..
Миофибриллы состоят из тонких актиновых и толстых миозиновых нитей.
Dмиоз=15нм, l=1,5мкм
В составе миозиновых нитей- белок миозин
Dакт=7нм, l=1мкм
В составе актиновых нитей:
-Актин ( F-актин) шаг спирали(?)≈38нм, 2 нити
- Тропомиозин 2-α-цепочки l≈41нм, шиг спирали ≈7нм
Функция тропомиозина – на нем акт. центры связывания с мостиками толстых нитей.
- Тропонин ( глоб. белок)- 3 суюъединицы- 2 молекулы на 1-м шаге
Миозиновая нить-( одна из самых больших структур в природе)
l≈155нм, d≈2нм( для молекул миозина)
Выделяют:
лёгкий меромиозин ( фибриллярный участок) l ≈110нм
миозиновый мостик обеспечивает скольжение нитей друг относительно друга
Тяжёлый меромиозин l≈45нм
Структура Миофибриллы

Уч. А – в центре светлая зона Н- без мостиков, не перекрытая тонкими нитями
I – изотропный участок
Все мостики на одной миозиновой нити работают синхронно.
Зона I в процессе сокращения изменяет свою длину (уменьшение), почти исчезает область Н.
23. Феноменологические соотношения между нагрузкой, скоростью сокращения и общей мощностью мышцы, эмпирические уравнения Хилла.

Где Q=a*x
E=Aактив+Qукор+ А±Qр
Х-укорочение мышцы
[a]=[P]=H
(P+a)*(V+b)=(P0+a)*b|V=0, Po max=a*(Vmax+b)|P=0, V max
PV+aV+Pb+ab=P0b+ab
 -
скорость, которую развивает мышца, в
изотоническом режиме ( в выражении для
мощности)
-
скорость, которую развивает мышца, в
изотоническом режиме ( в выражении для
мощности)
V max= b*P0/a ( при Р=0)
Nобщ=(Р+а)*V=b*(P0-P)
[b]=[V]=см/с (м/с)

а,b – некие характеристики отдельно взятой мышцы ( для каждой свои)
а/Р0 =0,25
хилл: а= f(P)=0,16*P0+0,18*P
24. Упругие свойства мышцы, режимы сокращения и их характеристики, понятие тетануса.
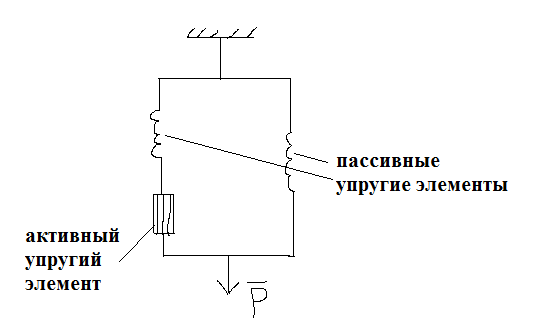
Упругие свойства мышцы имеют следующую структурную основу.
Компоненты:
- поперечные мостики,
- z-диски саркомеров,
- участки крепления миофибрина к сухожилиям,
- наружные элементы мышцы – коллагеновые элементы, сухожилия и др.
- места крепления мышц к костям,
- саркоплазматический ретикулум,
- сарколемма мышечного волокна.
Упругие компоненты
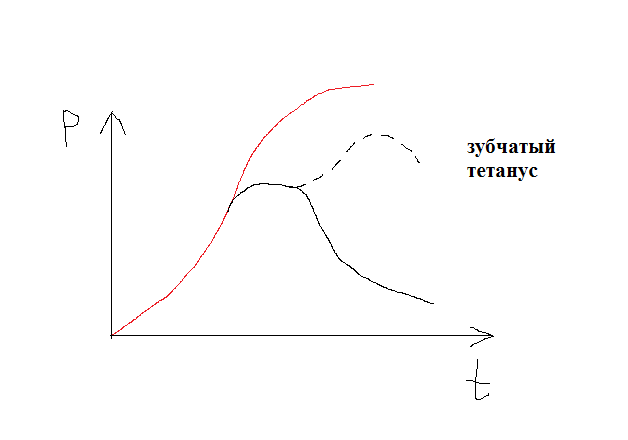
если 20-50 имп/сек -> гладкий тетанус
1/3 Ампл. – на одиночную стимуляцию
Меньше состыковок -> меньше развиваемое напряжение
Тетанус - длительное сокращение мышц, возникающее при последовательном воздействии на них ряда нервных импульсов, разделённых малыми интервалами.
Связь длина-напряжение при изометрическом сокращении
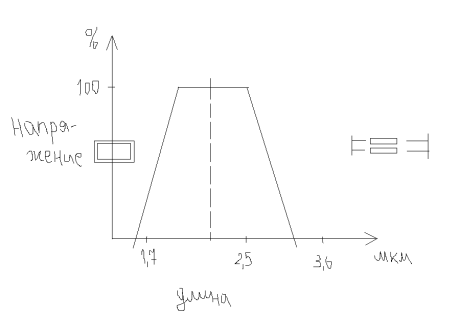
на участке 2..2,5 – плато – максимально развиваемое напряжение
Существуют два режима сокращения мышцы:
- изототическое сокращение (меняется длина волн, напряжение остается неизменным),
- изометрическое сокращение (концы мышцы жестко закреплены -> длина мышцы не меняется, меняется напряжение).
Если выполняются 2 сокращения:
- аоксотомическое сокращение.