

6. Моторная бляшка. Электронная микрофотограмма. 33 000.
1 - концевые веточки нервного волокна; 2 - митохондрии в аксоплазме; 3 - синаптические пузырьки в аксоплазме; 4 - аксолемма, образующая в этом месте пресинаптическую мембрану; 5 - сарколемма, образующая в этом месте постсинаптическую мембрану; 6 - складки постсинаптической мембраны; 7 - синаптическая щель; 8 - леммоцит (шванновская клетка); 9 - саркоплазма и 10 - ядро мышечного волокна (по Г. Еляковой). КОММЕНТАРИИ К ЭЛЕКТРОНОГРАММЕ
Моторная бляшка (нейромышечный синапс) - эффекторное (двигательное) нервное окончание, кото-рое встречается только в скелетной мышечной ткани. Нервное волокно (аксон + леммоцит) контактирует с миосимпластом.
Строение моторной бляшки похоже на строение классического синапса в нервной ткани. Поэтому видны следующие структуры:
1. Леммоцит (шванновская клетка) (цифра 8) - "прикрывает контакт сверху, изолируя и защищая его. В его цитоплазме видны митохондрии и цистерны гранулярной ЭПС.
2. Аксон двигательного нейрона (из передних рогов спинного мозга) (цифра 1), возле моторной бляшки уже не имеет миелиновой оболочки. Его аксолемма (цитолемма) (цифра 4) выполняет роль пресинаптической части синапса, поэтому в его аксоплазме много синаптических пузырьков (цифра 3), содержащих ацетилхолин (является медиатором в моторной бляшке). Кроме того имеются митохондрии, обеспечивающие энергию для транспорта медиатора из тела нейрона и его обратного захвата из синаптической щели.
3. Миосимпласт (мышечное волокна) в области моторной бляшки теряет поперечную исчерченность. В данном случае видны одно из его многочисленных ядер (цифра 10) и саркоплазма (цифра 9) - его сарколемма (цифра 5) выполняет роль постсинаптической мембраны и образует многочисленные складки в области синапса, для увеличения площади контакта с медиатором (цифра 6).
4. Синаптическая щель (цифра 7). КРАТКАЯ ИНФОРМАЦИЯ
Аксон мотонейрона выделяет ацетилхолин в синаптическую щель ==> рецепторы на постсинаптической мембране (в данном случае - сарколемме) возбуждаются ==> происходит деполяризация сарколеммы ==> волна деполяризации заходит в Т-трубочки (поперечные впячивания сарколеммы). Т-трубочки объединены в триады с концевыми цистернами (последние - это части саркоплазматического ретикулума, где депонируется Са++), следовательно, ==> возбуждение передается и на ретикулум, кальций выходит в саркоплазму ==> связывается с тонкими нитями, освобождая их активные центра для контакта с актином. Начинается сокращение миосимпласта.
Когда импульс перестает "приходить" по аксону, оставшийся в синаптической щели ацетилхолин разрушается ферментом ацетилхолинэстеразой или захватывается аксоном.
В скелетной мышечной ткани каждый симпласт имеет нейромышечный синапс, в отличие от гладкой и сердечной мышечных тканей, где один аксон просто ветвится среди 8-10 миоцитов. Нервное окончание в этом случае простое, свободное. Возбуждение между миоцитами передается по нексусам.
1 миосимпласт + 1 нервное волокно, образующее в ним синапс + капилляры вокруг симпласта (в составе эндонев-рия) = мион.
7.
Рецепторные (чувствительные) нервные окончания |
Эти нервые окончания - рецепторы - рассеяны по всему организму и воспринимают различные раздражения как из внешней среды, так и от внутренних органов. Соответственно выделяют две большие группы рецепторов:
Экстерорецепторы - стимулируемые окружающей средой
контактцепторы, воспринимающие раздражения, наносимые извне, и падающие непосредственно на ткани организма (болевые, температурные, тактильные и др.)
дистантцепторы, воспринимающие раздражения от источников, которые находятся на расстоянии (свет, звук)
Интерорецепторы
проприоцепторы, воспринимающие раздражения, возникающие внутри организма, в его глубоких тканях, связанных с функцией сохранения положения тела при движениях. Данный вид рецепторов представлен в мышцах, сухожилиях, связках, суставах, надкостнице, импульсы возникают в связи с изменением степени натяжения сухожилий, напряжения мышц и ориентируют в отношении положения тела и частей его в пространстве: отсюда еще наименование - "суставно-мышечное чувство", или "чувство положения и движения (кинестетическое чувство)".
висцерорецепторы воспринимающие раздражения от внутренних органов. Обычно от данных рецепторов информация очень редко доходит до сознания, например информация от барорецепторов, расположенных в каротидном синусе, которые непрерывно контролируют артериальное давление
В клинике приобрела довольно широкое распространение другая классификация, основанная на биологических данных [показать] .
С точки зрения биологических данных чувствительные нервные окончания рассматривается как соотношение и взаимодействие двух систем.
Одна, более древняя, свойственная более примитивной нервной системе, служит для проведения и восприятия сильных, резких, угрожающих целости организма раздражений; сюда относятся грубые болевые и температурные раздражения, связанные с древним "чувствующим" органом - зрительным бугром. Данная система чувствительности носит название протопатической, витальной, ноцицептивноп, таламической.
Другая система связана всецело с корой головного мозга. Являясь более новой и совершенной, она служит для тонкого распознавания качества, характера, степени и локализации раздражения. Сюда относятся такие виды чувствительности, как осязание, определение положения и движения, формы, места нанесения раздражения, различение тонких температурных колебаний, качества боли и т.д. Наименование этой системы чувствительности - эпикритическая, гностическая, корковая.
Эпикритическая чувствительность как система более новая, корковая, якобы оказывает тормозящее влияние на старинную протопатическую подкорковую чувствительность. Предполагалось, что в норме чувствительная функция человека определяется сосуществованием обеих систем в их определенной взаимосвязи; при этом эпикритическая чувствительность вносит элементы точного различения и анализа.
Такое подразделение чувствительности на два отдельных вида вызывает ряд серьезных возражений. Малодоказательным является представление об их соотношении как низшей и высшей систем, о тормозной функции эпикритической по отношению к протопатической; трудно представить себе роль зрительного бугра как органа, "воспринимающего" отдельные виды чувствительности.
В целостном организме любой вид чувствительности связан с работой коры больших полушарий, ибо всякое ощущение как акт сознания немыслимо без участия высших отделов головного мозга. Вместе с тем не подлежит сомнению, что в сложной чувствительности человека, достигшей высокого совершенства в процессе развития, существует представительство и древних примитивных систем, связанных с действием подкорковых, стволовых, сегментарных аппаратов. При поражении, выключении одного из звеньев высокодифференцированной чувствительной системы, значение в которой зрительного бугра все же остается несомненным, мы получаем качественно совершенно иную функциональную систему со своеобразным извращением ощущений и восприятий.
Более употребительной в клинической практике является описательная классификация, основанная на различении вида раздражения и возникающего в связи с ним ощушения.
В зависимости от специфичности раздражения, воспринимаемого данным видом рецептора, все чувствительные окончания разделяют на
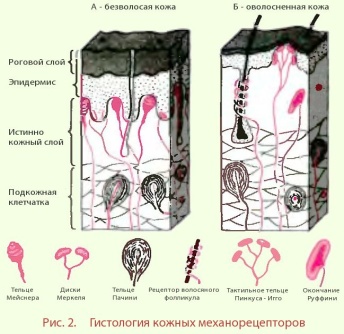
механорецепторы (рис.2)
медленно адаптирующиеся - например, давление веса тела на подошву. К ним относятся диск Меркеля - реагируют на деформацию перпендикулярно поверхности кожи, окончания Руффини (в безволосой коже) - реагируют на растяжение. В коже, покрытой волосами, диски Меркеля группируются под кожными возвышениями - тельцами Пинкуса-Игго.
быстро адаптирующиеся - реагирует только на механические стимулы, которые изменяются во времени. К ним относится тельце Мейснера (в безволосой коже), рецептор волосяного фолликула (в оволосненной коже)
очень быстро адаптирующиеся - реагируют на изменение скорости механической стимуляции. К ним относятся тельца Пачини. Тельца Пачини можно также назвать рецептором вибрации.
терморецепторы - температурная чувствительность (чувство холода и чувство тепла) и бессознательная регуляция температуры тела
барорецепторы - чувствительность к изменению артериального давления
хеморецепторы - чувствительность к понижению парциального давления кислорода и повышению углекислого газа, регулируют дыхание
ноцицепторы - чувство боли (поверхностной, глубокой, от внутренних органов)
рецепторы чувства позы, движения, мышечного усилия и др.
По особенностям строения чувствительные окончания подразделяются на
свободные нервные окончания, т. е. состоящие только из конечных ветвлений осевого цилиндра. Характерны для эпителия (холодовые рецепторы).
В этом случае миелиновые нервные волокна подходят к эпителиальному пласту, теряют миелин, а осевые цилиндры проникают в эпителий и распадаются там между клетками на тонкие терминальные ветви. В многослойном эпителии есть окончания, в состав которых входят, кроме терминалей отростков нервных клеток, специфически измененные эпителиальные клетки - осязательные эпителиоциты. От других клеток эпителия они отличаются светлой цитоплазмой, наличием осмиофильных гранул диаметром 65-180 нм и уплощенным темным ядром. Концевые нервные веточки подходят к таким клеткам и расширяются, образуя дисковидные концевые структуры, связанные с основаниями осязательных эпителиоцитов.
несвободные нервные окончания - содержащие в своем составе все компоненты нервного волокна, а именно ветвления осевого цилиндра и клетки глии
неинкапсулированныe - не имеющие соединительнотканной капсулы
инкапсулированныe - покрыты соединительнотканной капсулой. К ним относят пластинчатые тельца - рецепторы соединительной ткани, воспринимающие давление; осязательные тельца, находящиеся в составе сосочков кожи (тепловые рецепторы); мышечные веретена - рецепторы скелетных мышц и сухожилий, регистрирующие изменения длины мышечных волокон и скорость этих изменений; нервно-сухожильные веретена - так же рецепторы скелетных мышц и сухожилий, реагирующие на напряжение, прилагаемое к сухожилию при сокращении мышц.
Веретено состоит из нескольких (до 10-12) тонких и коротких поперечнополосатых мышечных волокон, заключенных в растяжимую соединительнотканную капсулу, - интрафузальных волокон.
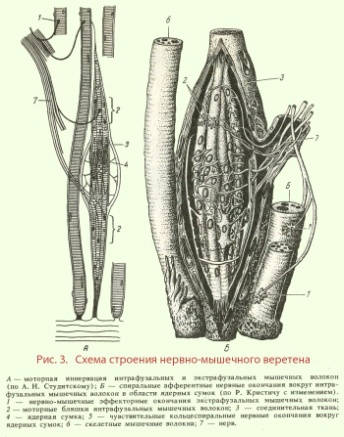
Остальные волокна мышцы лежат за пределами капсулы и называются экстрафузальными (рис. 3, А, Б). Интрафузальные волокна имеют актиновые и миозиновые миофиламенты только на концах, которые и сокращаются. Рецепторной частью интрафузального мышечного волокна является центральная, несокращающаяся часть.
Различают интрафузальные волокна двух типов: волокна с ядерной сумкой и волокна с ядерной цепочкой.
Волокон с ядерной сумкой в веретене 1-3. В центральной расширенной части они содержат много ядер.
Волокон с ядерной цепочкой в веретене может насчитываться 3-7. Они вдвое тоньше и вдвое короче, чем волокна с ядерной сумкой, и ядра в них расположены цепочкой по всей рецепторной области. К интрафузальным мышечным волокнам подходят афферентные волокна двух типов.
Первичные волокна диаметром 17 мкм образуют окончания в виде спирали - кольцеспиральные окончания как на волокнах с ядерной сумкой, так и на волокнах с ядерной цепочкой.
Вторичные волокна диаметром 8 мкм иннервируют волокна с ядерной цепочкой. По обеим сторонам от кольцеспирального окончания они образуют гроздъевидные окончания.
При расслаблении (или растяжении) мышцы увеличивается и длина интрафузальных волокон, что регистрируется рецепторами. Кольцеспиральные окончания реагируют на изменение длины мышечного волокна и на скорость этого изменения, гроздьевидные окончания - только на изменение длины. При внезапном растяжении из кольцеспиральных окончаний в спинной мозг поступает сильный сигнал, который вызывает резкое сокращение мышцы, с которой поступил сигнал - динамический рефлекс на растяжение. При медленном, длительном растяжении волокна возникает статический сигнал на растяжение, передаваемый как от кольцеспиральных, так и от гроздьевидных рецепторов. Этот сигнал может поддерживать мышцу в состоянии сокращения в течение нескольких часов.
Интрафузальные волокна имеют также эфферентную иннервацию. К ним подходят тонкие моторные волокна, оканчивающиеся аксомышечным синапсом на концах мышечного волокна. Вызывая сокращение концевых участков интрафузального волокна, они усиливают растяжение центральной рецепторной его части, повышая реакцию рецептора.
Нервно-сухожильные веретена обычно располагаются в месте соединения мышцы с сухожилием. Коллагеновые пучки сухожилия, связанные с 10-15 мышечными волокнами, окружены соединительнотканной капсулой. К нервно-сухожильному веретену подходит толстое (диаметром около 16 мкм) миелиновое волокно, которое теряет миелин и образует терминали, ветвящиеся между пучками коллагеновых волокон сухожилия. Сигнал с нервно-сухожильных веретен, вызванный напряжением мышцы, возбуждает тормозные нейроны спинного мозга. Последние тормозят соответствующие двигательные нейроны, предотвращая перерастяжение мышцы.
