

6. Фермент синтеза гликогена гликогенсинтетаза "а" (активная форма ), под воздействием цАмф-зависимой протеинкиназы переходит в неактивную гликогенсинтетазу "b".
Донором глюкозных остатков для синтеза гликогена служит УДФ-глюкоза, которая образуется из УТФ и глюкозо-1-фосфата под воздействием фермента глюкозо-1-фосфат-уридилтрансферазы:
глюкозо-1-фосфат + УТФ ® УДФ-глюкоза + Н3РО4.
Далее, под воздействием гликогенсинтетазы (1,4-гликозилтрансферазы) происходит перенос остатков глюкозы с УДФ-глюкозы на олигосахаридный фрагмент (гликоген - затравка).
Синтезируется линейная цепь гликогена, где остатки глюкоза соединяются друг с другом 1-4 альфа-гликозидной связью:
УДФ-глюкоза + n(глюкоза) ® УДФ + (1,4)-гликоген
Ветвления в молекуле гликогена возникают в результате действия фермента ветвления:
- амило-1,4-->1,6 гликозилтрансферазы.
(1,4)-гликоген (1,4)-(1,6)-гликоген
Гликоген — главная форма запасания углеводов у животных и
7. Гпиколипиды (3) содержатся во всех тканях, главным образом в наружном липидном слое плазматических мембран. Гликолипиды построены из сфингозина, остатка жирной кислоты и олигосахарида. Заметим, что в них отсутствует фосфатная группа. К наиболее простым представителям этой группы веществ относятся галактозилцерамид и глюкозилцерамид (так называемые цереброзиды). Соединения с сульфогруппой на углеводных остатках носят название сульфатидов.
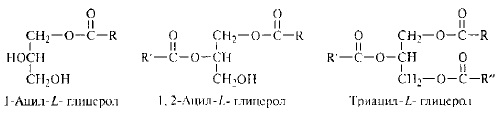
Гликолипиды участвуют также в иммунных реакциях, активном транспорте ионов натрия, передаче нервного импульса.
Билет33
1.Главной целью биологической химии является формирование системных знаний о закономерностях и химическом строении основных веществ организма и молекулярных основах биохимических процессов, лежащих в основе жизнедеятельности организма в норме и патологии.
2. После установления химической природы наследственного материала проблема самовоспроизведения (репликации) хромосом, а точнее — генотипа превратилась в проблему репликации ДНК. Первостепенное значение для решения этой проблемы имела разработка модели строения ДНК Ф. Криком и Дж. Уотсоном в 1953 г. Структура двойной спирали позволяла представить простой механизм репликации ДНК: двойная спираль сначала раскручивается, цепи расходятся, а затем каждая одноцепочечная половина молекулы ДНК достраивается до целой, двухцепочечной молекулы:Последовательность нуклеотидов вновь синтезирующихся цепей определяется правилом комплементарности оснований и последовательностью нуклеотидов имеющейся цепи. Иначе говоря, имеющиеся нуклеотидные цепи служат матрицей для синтеза новых цепей; в результате получаются две двухцепочечные молекулы ДНК, идентичные исходной молекуле. Такой способ репликации получил название полуконсервативного (в принципе возможен и другой механизм — консервативный, при котором вновь синтезируемая нуклеотидная цепь образуется прямо на двойной спирали ДНК, без ее раскручивания). Полуконсервативный механизм репликации ДНК нашел подтверждение в экспериментах с клетками кишечной палочки. Культуру Е. coli на протяжении нескольких поколений выращивали на среде, содержащей в качестве единственного источника азота 15NH4C1. После этого все вещества клеток Е. coli, в которые входит азот, содержали не обычный изотоп азота 14N, а тяжелый 15N. ДНК с 15N имеет большую плотность, чем ДНК с 14N, и это можно обнаружить методом центрифугирования (рис. 4.3). Если культуру, содержащую 151Ч-ДНК, пересеять на среду с немеченым азотом (14NH4C1), то ДНК клеток первого поколения имеет плотность, промежуточную между плотностями 15№ДНК и 14№ДНК; в клетках второго поколения обнаруживается два типа ДНК: с промежуточной плотностью и легкая (141Ч-ДНК). Эти результаты легко объясняются, если исходить из полуконсервативного механизма репликации ДНК (см. рис. 4.3). Позднее было установлено, что в клетках эукариот репликация ДНК происходит также полуконсервативным способом.
3. Ингибиторами ферментов называют вещества, снижающие их активность. Наибольший интерес представляют ингибиторы, взаимодействующие с активным центром фермента. Такие ингибиторы чаще всего являются структурными аналогами субстрата и, следовательно, комплементарны активному центру фермента. Поэтому они подавляют активность только одного фермента или группы ферментов с очень сходным устройством активного центра. Различают ингибиторы конкурентные и неконкурентные, обратимые и необратимые. Активаторы – вещества, которые повышают скорость ферментативных реакций, увеличивают активность ферментов. Они бывают органической и неорганической природы.
Активаторы органической природы: желчные кислоты (активируют поджелудочную ли пазу), энтерокиназа (активирует трипсиноген), глутатион, цистеин, витамин С (повышают активность оскидоредуктаз).
Активаторы неорганической природы: например, HCl активирует пепсиноген, ионы ме таллов (Na, Cl, K, Mg, Mn, Zn) активируют очень многие ферменты. Ионы металлов: а) спо собствуют образованию ферментсубстратного комплекса; б) служат донорами и акцептора ми электронов; в) принимают участие в образовании активного центра ферментов (Zn в со ставе карбангидразы, Fe – в составе цитохромов, каталазы, пероксидазы); г) выступают в ро ли аллостерических регуляторов.
4. Незаменимые аминокислоты — необходимые аминокислоты, которые не могут быть синтезированы в том или ином организме, в частности, в организме человека. Поэтому их поступление в организм с пищей необходимо.
Незаменимыми для взрослого здорового человека являются 8 аминокислот: валин, изолейцин, лейцин, лизин, метионин, треони́н, триптофан и фенилалани́н;
Для детей незаменимыми также являются аргинин и гистидин. полноценность белковых продуктов во многом определяется содержанием в них незаменимых аминокислот. Если в молекуле присутствуют все восемь незаменимых аминокислот, белки называются полноценными, они наиболее необходимы организму. Источники полноценных белков – это в основном продукты животного происхождения: молоко, яйца, рыба, мясо. Особенно ценны три незаменимые аминокислоты – триптофан, лизин, метионин. Повышенное количество триптофана содержится в молоке, мясе, зерновых продуктах; лизина – в мясе и продуктах из крови убойных животных, рыбе, твороге, яйцах; метионина – в твороге, мясе, горохе, фасоли, сое.
5. ХЕМИОСМОТИЧЕСКАЯ ТЕОРИЯ учение о механизме преобразования энергии в биол. мембранах при синтезе аденозинтрифосфорной к-ты (АТФ). Разработана П. Митчеллом в 1961—66. Согласно исходным представлениям Митчелла, запасание энергии в АТФ происходит вследствие предварительного, накопления зарядов на стенках мембраны, создания мембранного потенциала и разности концентраций протонов. Разность электрохимических. потенциалов ионов водорода на сопрягающих мембранах (внутренней мембраны митохондрий, тилакоиды хлоропластов, мембраны бактерий) возникает за счёт энергии, выделяемой при деятельности цепи окислит.-восстановит, ферментов, или за счёт поглощённых квантов света. Трансмембранные электрохимич. потенциалы ионов могут служить источником энергии не только для синтеза АТФ, на и для транспорта веществ, движения бактериальных клеток и др. энергозависимых процессов.
6. В организме человека и животных при недостаточном поступлении кислорода в клетки возможен, так называемый, анаэробный дихотомический путь катаболизма глюкозы (анаэробный гликолиз).
В интенсивно работающих скелетных мышцах, когда мощность механизма доставки кислорода в клетки оказываются недостаточными,
включаются анаэробные механизмы синтеза АТФ происходящие вне митохондрий и без участия митохондриальных ферментов.
В эритроцитах, где вообще отсутствуют митохондрии, потребность в АТФ удовлетворяется только за счет анаэробного гликолиза.
Интенсивный анаэробный гликолиз характерен и для опухолевых клеток.
7. Гиперлипопротеинемия - общее наименование пяти различных типов нарушений обмена веществ, при которых в крови больных обнаруживают жиры. Заболевание может быть наследственным. При некоторых формах гиперлипопротеинемий симптомы выражены слабо, от них можно избавиться, соблюдая диету. Тяжелое течение заболевания может привести к смерти.
Гиперлипопротеинемия обнаруживается примерно у каждого пятого человека с повышенным содержанием липидов и липопротеинов в крови. Это нарушение может быть также связано с наличием другого заболевания, например, диабета, болезни почек или поджелудочной и щитовидной желез.
Билет37
Роль биохимии в медицинской практике
Практически любое заболевание начинается с повреждения (нарушения) одной реакции в метаболизме клетки, а затем оно распространяется на ткань, орган и целый организм. Нарушение метаболизма ведет к нарушению гомеостаза в биологических жидкостях организма человека, что сопровождается изменением биохимических показателей.
Большое значение клинико-биохимических методов исследования биологических жидкостей велико в медицине и важно для подготовки медицинских лабораторных техников. Достаточно напомнить, что только в крови человека можно определить современными методами биохимических исследований около 1000 показателей метаболизма.
Вторичная структура белка. Связи, стабилизирующие вторичную структуру белка
В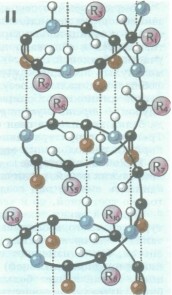 торичная
структура —полипептидная нить
закручена в спираль и образуются
водородные связи между СООН одного
виткаи NH2 другого витка, образуя
достаточно прочную структуру. Водородная,
пептидная связи
торичная
структура —полипептидная нить
закручена в спираль и образуются
водородные связи между СООН одного
виткаи NH2 другого витка, образуя
достаточно прочную структуру. Водородная,
пептидная связи
Коферменты ароматического ряда
Убихинон Химическое строение: Убихинон имеет бензольное кольцо, связанное с различными группами (2, 3-диметокси-метил-1,4-бензохинон с изопреновой цепью в шестом положении):
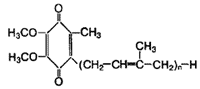
Убихинон — очень важный кофермент процессов биологического окисления питательных веществ и образования энергии в клетках. Входя в состав компонентов дыхательной цепи в митохондриях, он осуществляет перенос водорода через мембраны к цитохромам. Кофермент Q включен во многие пищевые смеси, которые используются для коррекции массы тела, повышения физической работоспособности, а также в растирочные препараты для улучшения энергообразования в суставах и мышцах.
Убихинон — весьма распространенный кофермент, отсюда его название — "вездесущий хинон". Он обнаружен во всех живых клетках. Внутри клеток убихинон локализован, по-видимому, только в митохондриях.
Строение , пищевые источники и биологические функции витамина С
А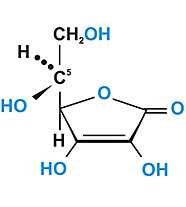 СКОРБИНОВАЯ
КИСЛОТА , С6Н8О6, водорастворимый
витамин. Синтезируется растениями (из
галактозы), животными (из глюкозы), за
исключением человека и приматов и
некоторых других животных, которые
получают аскорбиновую кислоту с пищей.
Биологическая роль аскорбиновой кислоты
связана с участием в окислительно-восстановительных
процессах клеточного дыхания.
СКОРБИНОВАЯ
КИСЛОТА , С6Н8О6, водорастворимый
витамин. Синтезируется растениями (из
галактозы), животными (из глюкозы), за
исключением человека и приматов и
некоторых других животных, которые
получают аскорбиновую кислоту с пищей.
Биологическая роль аскорбиновой кислоты
связана с участием в окислительно-восстановительных
процессах клеточного дыхания.
Влияет на различные функции организма: проницаемость капилляров, рост и развитие костной ткани, повышает иммунобиологическую сопротивляемость к неблагоприятным воздействиям, стимулирует продукцию гормонов надпочечников, способствует регенерации. Отсутствие аскорбиновой кислоты в пище человека понижает сопротивляемость к заболеваниям, вызывает цингу, или скорбут (язва во рту).
Суточная потребность в аскорбиновой кислоте составляет 60 мг — величина, не характерная для витаминов
Общие пути катаболизма. Окислительное декарбоксилирование альфа-кетоглутарата
- общие пути катаболизма - единые пути катаболизма для различных классов веществ, являющиеся продолжением специфических путей. С общими путями катаболизма, в основном, связаны главные биоэнергетические процессы, сопряженные с высвобождением и накоплением энергии в клетке.

Химизм и биологическое значение процесса резервирования гликогена
Механизм синтеза жиров в жировой ткани
В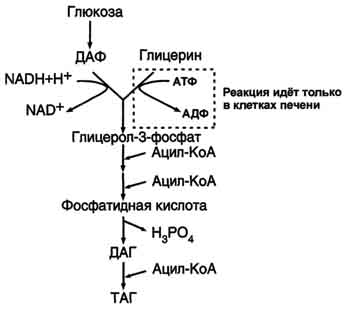 жировой ткани для синтеза жиров
используются в основном жирные кислоты,
освободившиеся при гидролизе жиров ХМ
и ЛПОНП (рис. 8-22). Жирные кислоты поступают
в адипоциты, превращаются в производные
КоА и взаимодействуют с глицерол-3-фосфатом,
образуя сначала лизофосфатидную кислоту,
а затем фосфатидную. Фосфатидная кислота
после дефосфорилирования превращается
в диацилглицерол, который ацилируется
с образованием триацилглицерола.
жировой ткани для синтеза жиров
используются в основном жирные кислоты,
освободившиеся при гидролизе жиров ХМ
и ЛПОНП (рис. 8-22). Жирные кислоты поступают
в адипоциты, превращаются в производные
КоА и взаимодействуют с глицерол-3-фосфатом,
образуя сначала лизофосфатидную кислоту,
а затем фосфатидную. Фосфатидная кислота
после дефосфорилирования превращается
в диацилглицерол, который ацилируется
с образованием триацилглицерола.
Кроме жирных кислот, поступающих в адипоциты из крови, в этих клетках идёт и синтез жирных кислот из продуктов распада глюкозы. В адипоцитах для обеспечения реакций синтеза жира распад глюкозы идёт по двум путям: гликолиз, обеспечивающий образование глицерол-3-фосфата и ацетил-КоА, и пентозофосфатный путь, окислительные реакции которого обеспечивают образование NADPH, служащего донором водорода в реакциях синтеза жирных кислот.
Молекулы жиров в адипоцитах объединяются в крупные жировые капли, не содержащие воды, и поэтому являются наиболее компактной формой хранения топливных молекул. Подсчитано, что, если бы энергия, запасаемая в жирах, хранилась в форме сильно гидратированных молекул гликогена, то масса тела человека увеличилась бы на 14-15 кг.
392
Билет38