

Билет27
1) Якоб Хендрик Вант-Гофф(1901) -в знак признания огромной важности открытия законов химической динамики и осмотического давления в растворах.
Фишер(1902)- за эксперименты по синтезу веществ с сахаридными и пуриновыми группами
Бухнер(1907)-за проведённую научно-исследовательскую работу по биологической химии и открытие внеклеточной ферментации
Вальтер Герман Нернст (присуждена в 1921 г.)-В признание его работ по термодинамике
Отто Виланд (присуждена в 1928 г.) -присуждена Нобелевская премия за исследования строения жёлчных кислот и аналогичных соединений
Уолтер Норман Хоуорс(1937)-за исследования углеводов и витамина С
Макс Фердинанд Перуц ,Джон Кодери Кендрю(1962)-за исследования структуры глобулярных белков
Уолтер Гилберт ,Фредерик Сенгер (1980)-за фундаментальные исследования биохимических свойств нуклеиновых кислот, в особенности рекомбинантных ДНК
Ханс Адольф КРЕБС- внёс основной вклад в разработку цикла трикарбоновых кислот (цикл Кребса). В 1932 описал орнитиновый цикл синтеза мочевины в печени животных.Лауреат Нобелевской премии по медицине в 1953 г. совместно с Фрицем Липманом.
2) Генетический код для аминокислот является вырожденным. Это означает, что значительное большинство аминокислот кодируется несколькими кодонами. За исключением метионина и триптофана, Узнавание кодона мРНК антикодоном тРНК основано не только на спаривании оснований, когда каждое основание кодона образует пару оснований с комплементарным азотистым основанием антикодона. В этом случае каждый антикодон, соответственно каждая молекула тРНК, может в принципе узнавать только один кодон мРНК. Вырожденность генетического кода имеет биологический смысл, обеспечивая организму ряд преимуществ. В частности, она способствует «совершенствованию» генома, так как в процессе точечной мутации, вызванной химическими или физическими факторами, возможны различные аминокислотные замены. Другой отличительной особенностью генетического кода является его непрерывность, отсутствие «знаков препинания», т.е. сигналов, указывающих на конец одного кодона и начало другого. Другими словами, код является линейным, однонаправленным и непрерывающимся: АЦГУЦГАЦЦ. Это свойство генетического кода обеспечивает синтез точной и в высшей степени упорядоченной последовательности аминокислотных остатков в молекуле белка.
3) Лактатдегидрогеназа (ЛДГ) – фермент, обратимо катализирующий окисление лактата в пируват. Известны 5 изоформ ЛДГ, из них большее диагностическое значение имеют ЛДГ1 и ЛДГ5. ЛДГ1 участвует в окислении лактата в пируват и преобладает в тканях с аэробным типом метаболизма (миокард, почки, мозг, эритроциты, тромбоциты). ЛДГ5 напротив, способствует превращению пирувата в лактат и активна в тканях с высоким уровнем гликолиза (печень, скелетные мышцы). Основная роль общей ЛДГ заключается в выявлении тканевого повреждения. Выявлена высокая специфическая активность для печени, миокарда, скелетной мускулатуры, почек и эритроцитов. При мышечной дистрофии Дюшенна увеличение активности ЛДГ выявляется за несколько лет до обнаружения клинических сиптомов. Определение активности изоферментов ЛДГ1 более информативно при заболеваниях крови, почек, миокарда, а ЛДГ5 – при заболеваниях гепатобилиарной системы. Активность ЛДГ также повышается при циррозе, опухолях различной локализации, травмах, лекарственной интоксикации
4) Биоти́н — водорастворимый витамин группы В. Молекула биотина состоит из тетрагидроимидазольного и тетрагидротиофенового кольца, в тетрагидротиофеновом кольце один из атомов водорода замещен на валериановую кислоту.
содержится в говядине, телятине, курином мясе и ветчине, коровьем молоке и сыре, сельди. Из растительных продуктов им богаты помидоры, соевые бобы, неочищенный рис и рисовые отруби, пшеничная мука, морковь и цветная капуста, яблоки и апельсины, бананы и дыня, картофель, свежий лук, цельные зёрна ржи. В тканях животных биотин связан с белками, а во фруктах и овощах он содержится в свободном состоянии. Биотин занимает важную роль в процессах обмена белков, жиров и углеводов, необходим для активации витамина С, с его участием протекают реакции активирования и переноса углекислого газа в кровеносной системе, формирует часть некоторых ферментных комплексов и необходим для нормализации роста и функций организма. Биотин, взаимодействуя с гормоном инсулином, стабилизирует содержание сахара в крови, также участвует в производстве глюкокиназы. Оба этих фактора важны при диабете. Рекомендуемой суточной дозой биотина является:
для взрослых 150-300 мкг для детей, в зависимости от возраста и пола 50-150 мкг
5) АТФ – это универсальное энергетическое вещество организма. АТФ – нуклеотид, в состав молекулы которого входят азотистое основание – аденин, углевод – рибоза и три остатка фосфорной кислоты. Особенностью молекулы АТФ является то, что второй и третий остатки фосфорной кислоты присоединяются связью, богатой энергией, иначе называемой макроэргической связью. Часто соединения, имеющие макроэргическую связь (а мы столкнемся с ними в процессе изучения предмета) обозначатся термином «макроэрги» или макроэргические вещества.
Строение АТФ:
^ Аденин – рибоза – Ф.К. – Ф.К. – Ф.К.
аденозин
При использовании АТФ в качестве источника энергии обычно происходит отщепление путем гидролиза последнего остатка фосфорной кислоты.
^ АТФ + Н2О → АДФ + Н3РО4 + энергия
Главными потребителями энергии АТФ в организме являются
реакции синтеза; мышечная деятельность; транспорт молекул и ионов через мембраны
6)
7 При смешанной диете примерно половина суточной нормы холестерина синтезируется в кишечнике, коже и главным образом в печени (примерно 50%), а остальной холестерин поступает с пищей. Значительная часть холестерина включена в липидный слой плазматических мембран. Большое количество холестерина расходуется в биосинтезе желчных кислот (см. с. 306), часть выделяется с желчью. Ежесуточно из организма выводится примерно 1 г холестерина. Очень небольшая часть холестерина используется для биосинтеза стероидных гормонов
Главным источником для синтеза холестерина является ацетил-КоА, и до стадии образования b-гидрокси-b-метилглутарил-КоА (ГМГ-КоА)
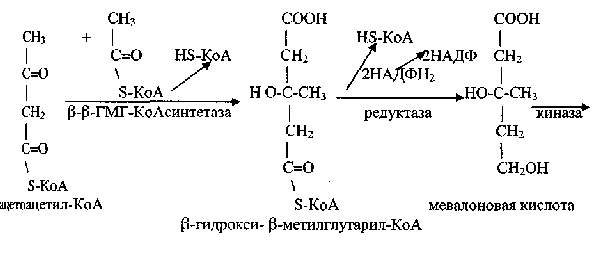
П од воздействием мевалонат киназы с использованием двух молекул АТФ образуется мевалонилпирофосфат, при декарбоксилировании которого образуется изопентилпирофосфат и далее диметилаллилпирофосфат:
Билет28
1)Биохимия (биологическая химия) — наука о химических процессах, происходящих в живых организмах. Главной задачей, является изучение механизмов и закономерностей, лежащих в основе процессов формирующих и поддерживающих живое состояние.
2) Так как транскрипция связывает ядро –клетки, и белки клетки, то от качества и активности транскрипции зависит объем синтеза тех или иных белков, жизнедеятельность клетки, ее способность адаптироваться к окружающей обстановке.
Транскрипция- это копирование генет. инф-ии, заключенной в генах молекулы ДНК. С образованием молекул м-РНК.
Гены молекулы ДНК, принимающие участие в синтезе РНК объединены в отдельные функциональные группы, получившие название оперон. Такая функциональная организация позволяет эффективнее регулировать экспрессию (транскрипцию) этих генов.
Оперонная модель регуляции экспрессии генов предполагает наличие единой системы регуляции у таких объединенных в один оперон структурных генов, имеющих общий промотор и оператор.
3) НАД и НАДФ являются производными витамина РР (В5. никотинамид) и служат в качестве кофермента анаэробных дегидрогеназ: изоцитрат дегидрогеназа, лактатдегидрогеназа, малатдегидрогеназа, оксиацил КоА-дегидрогеназы.
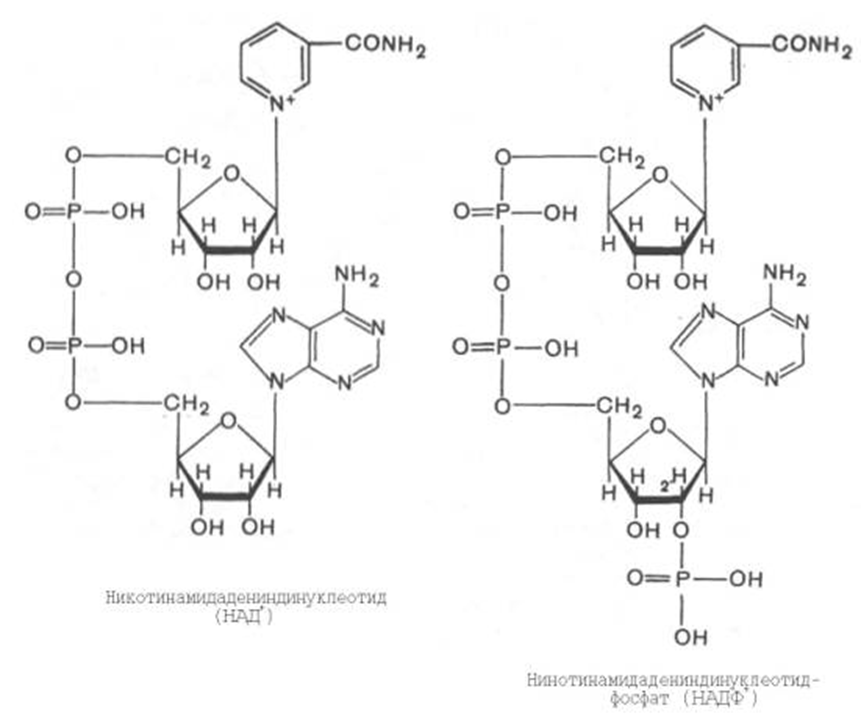
Основной функцией биологического окисления является обеспечение организма энергией в доступной для использования форме (прежде всего в форме АТФ).
В переносе электронов от субстратов к молекулярному кислороду принимают участие:
1.Пиридинзависимые анаэробные дегидрогеназы, для которых коферментами служат либо НАД, либо НАДФ
Эти ферменты катализируют реакции переноса атомов водорода (протоны и электроны) на любой акцептор кроме кислорода
Представители:
Пируват дегидрогеназа
Альфа-кетоглутаратдегидрогеназа
Изоцитрат дегидрогеназа
Малат дегидрогеназа
Коферментом этих ферментов являются НАД, НАДФ- производные витамина РР ( В5- никотиновая кислота)
Аэробные дегидрогеназы
Эти ферменты катализируют реакции переноса атомов водорода (протоны и электроны) на любой акцептор, в том числе и на кислород
Представители:
сукцинатдегидродгеназа
глицерофосфатдегидрогеназа
ацил-КоА-дегидрогеназа
Коферментами этих ферментов являются ФАД, ФМН- производные витамина В2 ( рибофлавин)
4) Микроэлементы- химические элементы, присутствующие в тканях человека, животных и растений в так называемых малых количествах. Биологическая роль М. определяется их участием практически во всех видах обмена веществ организма; они являются кофакторами многих ферментов (Ферменты), витаминов (Витамины), гормонов, участвуют в процессах кроветворения, роста, размножения, обеспечивающих нормальную жизнедеятельность организма. Обеспеченность М. обусловлена их содержанием в воде и пищевых продуктах (Пищевые продукты), количественным соотношением М. между собой и усвояемостью тех веществ, в составе которых М. поступают в организм. Содержание М. в пищевых продуктах во многом зависит от геохимической зоны. Существуют районы со значительными отклонениями концентрации определенных М. в почве и воде от нормы (как в сторону уменьшения, так и увеличения), что отражается на содержании этих М. в продуктах растительного и животного происхождения.
5) Окислительное декарбоксилирование пирувата катализируется мультиэнзимным пируват-дегидрогеназным комплексом, локализованным на внутренней мембране митохондрий со стороны матрикса.
Пируват-дегидрогеназный комплекс (окислительная декарбоксилаза пировиноградной кислоты) состоит из трех ферментов (Е1,Е2,Е3):
Е1-пируватдекарбоксилаза с коферментом ТПФ (тиаминпирофосфат),
Е2-дигидролипоил-трансацетилаза с коферментом липоевая кислота (ЛК) и кофактором - НS-КоА,
Е3-дигидролипоил-дегидрогеназа с коферментом ФАД и кофактор - НАД

6) Гиалуроновая кислота – состоит из β,D-глюкуроновой кислоты и N-ацетилглюкозамина, которые связаны β-1,3-гликозидной связью. Дисахаридный фрагмент повторяется многократно и образует гиалуроновую кислоту.
К примеру, гиалуроновая кислота является важной составляющей суставного хряща, играющей роль биологической смазки. Эта смазка препятствует трению суставных поверхностей, предупреждая их преждевременное изнашивание.
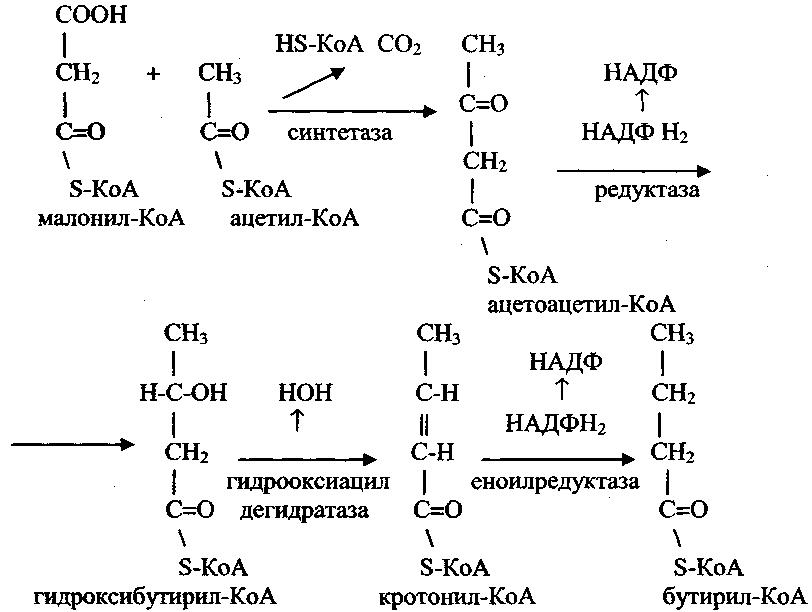
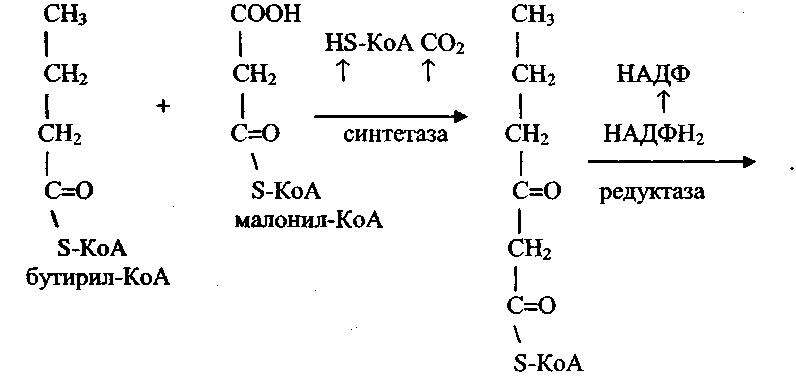
7) Биосинтез жирных кислот. Источником для синтеза жирных кислот служит малонил-КоА, образующийся из ацетил-КоА.
Ацетил-КоА доставляется к месту синтеза жирных кислот из митохондрий, где он образуется при бета-окислении жирных кислот.
Доставка ацетил-КоА из митохондрий в цитозоль осуществляется либо карнитиновым челночным механизмом, либо с помощью цитрата.
В митохондриях из ацетил-КоА и щавелево-уксусной кислоты (ЩУК) синтезируется цитрат, который при участии специфической транслоказы митохондриальной мембраны доставляется в цитозоль.
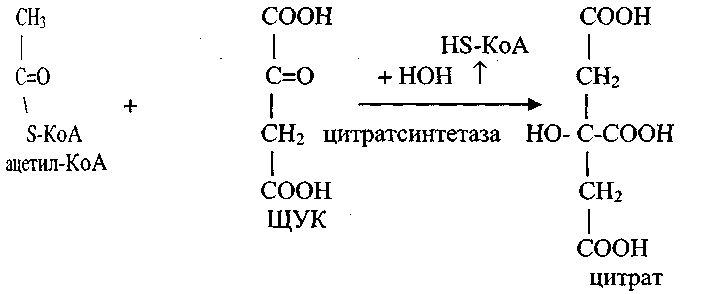

Билет29.
1)функциональная биохимия занимается анализом химических процессов, лежащих в основе определенных проявлений жизнедеятельности.
2) Различают 4 уровня структурной организации белка: первичную, вторичную, третичную и четвертичную.