

1) Предмет и задачи микологии. Краткие сведения по истории микологии. Традиционные и современные представления о положении грибов в системе органического мира Осмотрофное питание как основной признак грибов. Грибы и грибоподобные организмы – сходство и различия.
Грибы – большая группа эукариотных гетеротрофных организмов с осмотрофным типом питания и «наружным пищеварением», размножающаяся спорами, включающая от 100 тысяч до 250 тыс видов. Для большинства из них характерно:
-неподвижность в вегетативном состоянии
- неограниченный рост вегетативного тела
- наличие клеточной стенки.
1745 год Карл Линней отнес грибы к царству растений.
19 век – предложения выделить грибы в самостоятельное царство.
Во второй половине 20 века наблюдалось бурное развитие физиологии и биохимии. Новые данные окончательно убедили научное сообщество, что грибы нельзя отнести к царству растений. Их отличия от растений: грибы – это гетеротрофные редуценты, оболочки клеток хитиновые, отличен углеродный и азотный обмены (образуются гликоген, серотонин, мочевина), особый путь синтеза лизина, другой состав стеролов. Отличие от животной клетки Protozoa: осмотрофный (абсорбционный) способ питания (всей поверхностью тела), размножение спорами, неподвижность тела. Современные материалы по биохимии и физиологии, ультраструктуре клетки и строению клеточной оболочки грибов позволяют считать, что грибы занимают промежуточное положение между животными и растениями и позволяют предположить, что они представляют отдельную филогенетическую ветвь живой природы – царство Myxota, или Fungi. Наибольшее филогенетическое значение в систематике грибов придается наличию или отсутствию у них подвижности, строению подвижных стадий (зооспор, гамет) и составу клеточной стенки.
Два типа жгутиков: бичевидный гладкий и перистый. Если у клетки два одинаковых – изоморфные жгутики, разные – гетероконтные.
Сейчас грибы делят на две группы: грибоподобные организмы и настоящие грибы.
|
Грибы |
Грибоподобные организмы |
||
Клеточная стенка |
Хитин и глюкан |
Целлюлоза и глюкан |
||
митохондрии |
Кристы плоские |
Кристы трубчатые |
||
Подвижные клетки |
Жгутики отсутствуют или задние |
Жгутики передние и боковые, гетероконтные |
||
Запасной продукт |
Гликоген + мочевина и тригалоза (микоза) |
миколаминарин |
||
Биосинтез лизина |
|
|
||
стеролы |
эргостерол |
фукостерол |
||
Отделы |
Chytridiomycota Zygomycota Glomeromycota Ascomycota Basidiomycota |
Oomycota Myxomycota |
2) Вегетативное тело грибов (клетка с ризомицелием: моноцентрические и полицентрические формы, интраматрикальное и экстраматрикальное расположение таллома; плазмодий, мицелий, дрожжеподобные клетки). Мицелиально-дрожжевой диморфизм. Основные типы мицелия (септированный и ценотический; эндофитный и поверхностный; пигментированный и бесцветный). Рост мицелия, апикальный аппарат гиф. Видоизменения мицелия (анастомозы, пряжки, апрессории, гаустории, ловчие кольца, столоны, ризоиды, оидии, хламидоспоры). Сплетения мицелия (пленки, тяжи, ризоморфы, склероции). Понятие о ложных тканях.
Вегетативное тело грибов – таллом, или мицелий, представляет собой систему ветвящихся трубок, гиф, с апикальным ростом и боковым ветвлением.
Ценотический (неклеточный) мицелий лишен перегородок, т.е. это большая масса цитоплазмы, содержащая много ядер. Характерен для ряда представителей Chytridiomycota, Oomycota, Zygomycota.
Клеточный (септированный, компартментализированный) мицелий разделен септами (перегородками) на одно-, дву- или многоядерные клетки. Хар-ен для сумчатых, базидиальных и анаморфных грибов. При делении септа врастает с боков к центру. В центре обычно остается пора. Если септа перфорирована многочисленными микропорами – микропоровая септа ( Хитридиомикота). Одна центральная пора, причем септа уменьшается по направлению к поре – простая септа (сумчатые аскомикота). Септа утолщается по направлению к поре – долипоровая септа (базидиальные и высшие грибы). Причем у некоторых базидиальных пора имеет колпачок –парентосому.
Мицелий, пронизывающий субстрат, - субстратный (погруженный) мицелий.
Располагающийся на поверхности – поверхностный (воздушный) мицелий.
Существуют немицелиальные грибы. Например у паразитов (хитридиевые и гифохитридиевые) таллом одноклеточный, микроскопический, спорической или эллипсоидной формы. От клетки могут отходить тонкие разветвленные нитевидные структуры – ризомицелий, пронизывающий субстрат. Одна клетка-центр образования ризомицелия и репродуктивных органов – моноцентрический ризомицелий. Несколько центров – полицентрический.
У одноклеточных видов без ризомицелия все вегетативное тело идет на формирование репродуктивных органов – холокарпические виды. Виды, имеющие ризомицелий, - эукарпические. Их таллом лежит внутри субстрата – интраматрикальные, на поверхности, погрузив в субстрат мицелий, - экстраматрикальные виды.
У дрожжей (сумчатые грибы) вегетативное тело представлено одиночными почкующимися клетками. Если они не расходятся, образуется псевдомицелий.
Видоизменения мицелия и сплетения гиф:
- Апрессории или присоски
- Гаустории: мицелий своим кончиком проникает сквозь мембрану клетки хозяина, тихонько надавливая, образуется кармашек, туда залезает эта веточка мицелия, там начинает расширяться – капсула Гаустория. Гриб забирает все активные соединения клетки.
- ловчие кольца: ловят личинок нематод, внутрь тела нематоды врастают питающие нити, высасывающие органику.
- столоны (воздушные гифы)
- ризоиды (прикрепление к субстрату)
- оидии - отдельные клетки мицелия. На мицелии образуются перегородки, формируются короткие фрагменты, из перегородки исчезает пора, формируется перетяжка, мицелий распадается.
- склероции – прочные переплетения мицелия, образующие комочек. Обычно темноокрашенные. Служат для перенесения неблагоприятных условий. Разделяют на истинные и ложные. У истинных на поперечном срезе 2 слоя: узкий коровый (образован очень плотным сплетением мелких гиф с толстыми оболочками, пропитанными меланином) (прочность) и сердцевинный (сплетение крупных гиф с тонкими оболочками и запасом питательных веществ). Некоторые грибы образуют склероции, пронизывая и мумифицируя ткань хозяина – псевдосклероции.
- стромы – сплетения мицелия, защищающие плодовые тела сумчатых грибов.
- хламидоспоры – отдельные клетки мицелия, покрываются толстыми оболочками и обособляются. Увеличен запас питательных веществ и ядер.
- мицелиальные тяжи (шнуры) – состоят из гиф, расположенных параллельно, между которыми могут быть анастомозы. На м. тяжах в почве образуются плодовые тела шляпочных грибов. Проводящая функция.
- ризоморфы: наружные гифы утолщенные, темноокрашенные (защитная функция), внутренние – тонкостенные (проводящая функция). До нескольких метров в длину! Распространение грибы по субстрату.
- пленки – гифы плотно сплетаются, не срастаясь.
Ложные ткани: плектенхима – плотное переплетение гиф, из нее состоят плодовые тела шляпочных грибов.
Плазмодий – амебоидный таллом без клеточной стенки, много ядер.
3) Ультраструктурные особенности грибной клетки (клеточная стенка, строение пор: долипоры и простые поры аскомицетного типа +; особенности основных органелл: эндоплазматический ретикулюм, цистерны Гольджи, митохондрии, микротельца). Митоз у грибов (общие черты, особенности метафазы и анафазы).
Клеточная стенка: структура из хитина и целлюлозы. Иногда хитин заменен на хитозан и маннан. Субмембранный комплекс: к мембране приближены микрофиламенты (актин), почти нет микротрубочек (тубулин).
Цитоплазматическая мембрана – прижата к клеточной стенке, роль поддержания осмотического давления и определенного электрического потенциала. Ломосомы – выпячивания цитоплазматической месьраны со всевозможными пузырьками.
Основные органеллы:
Митохондрии,
ЭПС у грибов обычно гладкая, шероховатая представлена слабо, рибосомы свободные, рассеянные по цитоплазме,
аппарат Гольджи не выражен, есть цистерны Гольджи (по-видимому относятся к ЭПС), пузырьки отшнуровываются от любых частей цистерны, аппарат Гольджи только в зооспорах, у Oomycota иногда и в вегетативных клетках мицелия,
Вакуоль с клеточным соком, окруженная мембраной (тонопластом).
Ядра мелкие, одно или много.
Митоз у грибов (все типы ядер):
- закрытый или частично закрытый
- отсутствие центриолей
- отсутствие метафазной пластинки (особенность метафазы)
- асинхронное расхождение хромосом в анафазе
Хромосомы редко выстраиваются по экватору, следовательно, много хромосомных мутаций и активное видообразование. Те, в ядра которых попало меньше, чем нужно, хромосом, погибают. В которых больше – задействуются разные участки.
4) Размножение грибов. Разнообразие репродуктивных механизмов грибов. Вегетативное размножение (мицелий, оидии, столоны, почкование)+. Бесполое размножение (эндоспоры: зооспоры и спорангиоспоры и экзоспоры: конидии). Холокарпические и эукарпические (поликарпические) формы+. Разнообразие конидиеносцев и конидий. Организация конидиального аппарата (свободные конидиеносцы, коремии и конидиомы: ложе и пикниды). Роль бесполого размножения в жизненных циклах грибов. Биологический смысл перехода к конидиальному спороношению.
Вегетативное размножение: почкование дрожжей, оидии, столоны и ризоиды, хламидоспоры.
Бесполое размножение – анаморфа.
1. Эндоспоры – споры, внутри любой клетки, могут быть подвижны – зооспоры (у грибоподобных организмов, у грибов – только у хитридиевых, у всех остальных грибов – без жгутиков). Холокарпические формы – на зооспору идет все вегетативное тело, эукарпические – зооспорангий как отдельный орган. Спорангиоспоры – неподвижные, без жгутиков, образуются внутри специализированного спорангия (Zygomycota), на спорангиеносцах. Распространение спорангиоспор связано с разрывом спорангия (в результате механического воздействия или изменения влажности), что не очень удобно. Тенденция к уменьшению количества спор в спорангиях и утолщению облочки.
2. Экзоспоры – формируются открыто, на веточках мицелия – конидиях. Конидии очень распространены у грибов, клеточка на специальной веточке мицелия, по одной или цепочкой, формируется конидиеносец:
1) таллический. Участвует только одна веточка, в результате конидиеносец перестает существовать.
2) бластический. Есть конидиогенная клетка, дает конидии, не разрушается.
Конидиомы – скопление конидиеносцев, защищенных специальными образованиями. Коремии – конидиеносцы сближены.
Ложе: сплетение мицелия – основание ложа, на нем масса конидиеносцев, на которых образуются конидии. Прикрыто субстратом (погружено в него), пленочкой мицелия. Пикнида – полузамкнутое вместилище, количество конидий нарастает, конидиеносцы услизняются и выдавливают конидии из пикниды.
Сложная пикнида – строма. Сплетения с полостями, внутри них – конидии, глубоко погружена в субстрат.
Тенденция к утрате жгутиковых стадий.
5) Половое размножение: хологамия, гаметогамия (изогамия, гетерогамия, оогамия и ее особенности), гаметангиогамия (в том числе зигогамия), соматогамия, спермациогамия (=дейтерогамия). Гомо- и гетероталлизм. Особенности полового размножения сумчатых грибов (формы полового процесса, формирование сумки, разнообразие сумок). Особенности полового размножения базидиальных грибов (форма полового процесса, формирование базидии, разнообразие базидий, представление о базидиоспорах как о баллистоспорах).
Форма - Телеоморфа.
Существенные моменты полового процесса грибов: плазмогамия, кариогамия и мейоз. Гриб может находится в гаплоидной и диплоидной стадиях. В результате полового процесса образуются гаплоидные, неоднородные в генетическом отношении споры, они располагаются на мицелии, на поверхности или внутри плодовых тел, называемых телеоморфами. Т.е. половое размножение дает начало формам с новой комбинацией генетического материала.
У грибов известно три основных типа полового процесса: гаметогамия, гаметангиогамия и соматогамия.
Гаметогамия – слияние гамет, образующихся в гаметангиях. Изогамия и гетерогамия. Оогамия (яйцеклетки из оогоний, сперматозоиды из антеридий). У многиз грибов сперматозоиды не образуются, я-ка оплодотворяется недифференцированным содержимым выростов многоядерного антеридия. Хитридиомикота и гифохитридиомикота.
Гаметангиогамия – слияние двух многоядерных специализированных структур, содержимое которых не дифференцировано на гаметы. Хаар-на для зиго и аскомикот. У зигомикот – зигогамия: слияние многоядерных клеток, гаметангиев, не дифференцированных на мужской и женский. Из зиготы образуется одетая толстосетнной окрашенной оболочкой зигоспора, которая прорастает после периода покоя в особый зародышевый спорангий. У аскомикот половые органы дифференцированы на мужской и женский (антеридий и аскогон). Аскогон состоит из крупной многоядерной клетки (собственно аскогона) и тонкой нитевидной – трихогины, через которую в аскогон переливается содержание многоядерного антеридия. Далее из оплодотворенного аскогона вырастают аскогенные дикариотичные гифы. Ядра сливаются, диплоидные ядра в дальнейшем делятся мейотически. Формируются сумки, или аски, внутри митоз и формирование восьми гаплоидных аскоспор.
Соматогамия – сливаются обычные вегетативные клетки мицелия. Хитридиомикота и гифохитридиомикота., имеющие одноклеточный таллом. Хологамия – слияние одинаковых вегетативных клеток, являющихся самостоятельными организмами. У базидиомикот слияние двух вегет. клеток гаплоидного мицелия. Сначала плазмогамия и образуются дикарионы и дикариотический мицелий. Далее на этом мицелии образуются базидии, где происходит слияние ядер дикариона и мейоз, и на базидии формируются экзогенные гаплоидные базидиоспоры. При созревании они отбрасываются (гидролиз гликогена, повышение тургорного давления). Поэтому их еще называют баллистоспорами.
Т.е. эндогенные аскоспоры сумчатых грибов и экзогенные базидиоспоры базидиальных образуются в результате полового процесса.
У грибов различают гомоталические (обоеполые) и гетероталлические (раздельнополые) формы. У гетероталличных грибов на мицелии, выросшем из одной споры, половые органы не закладываются, они развиваются лишь при встрече двух мицелиев, отличающихся по половому знаку.
6) Плодовые тела: аскомы и базидиомы. Разнообразие плодовых тел сумчатых грибов (клейстотеции, перитеции, апотеции, асколокуляоные стромы = псевдотеции). Разнообразие плодовых тел базидиальных грибов. Базидиомы гимениального типа (строение гимения: базидиолы, цистиды, щетинки; разнообразие гименофоров). Афиллофороидные базидиомы: разнообразие и основные направления эволюции. Агарикоидные базидиомы: гимнокарпное и гемиангиокарпноес развитие; частное и общее покрывало. Базидиомы гастероидного типа: строение молодого и зрелого плодового тела; разнообразие способов распространения базидиоспор.
Отдел Ascomycota: плодовые тела – аскомы. Три типа:
Клейстотеций – замкнутое шаровидное тело, одетое плотной, чаще темноокрашенной оболочкой – перидием. Сумки в нем располагаются беспорядочно. Оболочки сумок прототуникатные, быстро разрушаются. Сумки и аскоспоры освобождаются пассивно после разрушения перидия и оболочек сумок.
Перитеций – полузамкнутое плодовое тело округлой, грушевидной или кувшинообразной формы с узким отверстием на вершине, через которое происходит активное выбрасывание аскоспор из сумок. Сумки унитуникатные, цилиндрические или булавовидные, расположены в перитеции пучком или слоем. Между ними находятся стерильные нити – парафизы. В верхней части перитеция – носике, обычно расположены перифизы – короткие нитевидные гифы, направленные к отверстию перитеция. Сумки поочередно подрастают к отверстию перитеция. Под действием тургорного давления их оболочки разрываются, а аскоспоры отбрасываются на расстояние до нескольких сантиметров. Затем оболочка этой сумки отпадает и на ее место вырастает другая сумка.
Апотеций – блюцевидное или чашевидное открытое полое плодовое тело. На его верхней стороне расположен слой, состоящий из унитуникатных сумок и парафиз, называемый гимением. Аскоспоры из сумок отбрасываются активно. Одновременно высвобождаются споры из многих сумок.
Асколокулярные стромы = псевдотеции. Сумки эутуникатные, образуются в особых полостях (локулах), возникающих в сплетении мицелия – аскостроме, или псевдотеции.
Отдел Basidiomycota. Плодовые тела – базидиомы. Очень разнообразны по форме, консистенции и окраске.
Спороносный слой плодового тела – гимений – у более примитивных располагается на верхней, а у более высокоорганизованных видов – на нижней стороне плодового тела.
Гимений состоит из базидий с базидиоспорами и базидиол (молодые или недоразвитые базидии). Также могут быть цистиды – крупные клетки, возвышающиеся над гимениальным слоем. Они защищают гимений и базидии от давления сверху.
Поверхность плодового тела, несущая гимений, называется гименофором. Может быть гладким, иметь форму зубцов, трубочек, пластинок. (гладкий, шиповатый, трубчатый, пластинчатый, лабиринто-сетчатый, бугорчатый гименофор).
Афиллофороидные гименомицеты:
Плодовые тела не дифференцированы на шляпку и ножку, гименофор гладкий, складчатый, зубчатый, трубчатый. Плодовые тала распростертые, блюцевидные, чашевидные, вееровидные и другие, гименофор всегда на нижней стороне. Можно предполагать эволюцию этих форм от слабо развитых, рыхлых плодовых тел с гладким гименофором на верхней стороне до хорошо развитых плодовых тел со сложным гименофором, обычно трубчатым, на нижней стороне.
Структура гиф в плодовых телах разная.
Генеративные гифы септированные, могут иметь пряжки. Дают начало базидиям.
Вегетативные гифы – нет септ. Делятся на связующие (тонкие, разветвленные) и скелетные (толстые, неразветвленные).
Структура плодовых тел:
- мономитическая – есть только генеративные гифы.
- димитическая – генеративные + связующие ИЛИ скелетные гифы
- тримитическая – все три типа.
Агарикоидные гименомицеты:
Мягкие, однолетние плодовые тела, дифференцированные на ножку и шляпку. На нижней стороне шляпки пластинчатый гименофор (только у болетовых трубчатый).
Если гименофор закладывается открыто – гимнокарповые плодовые тела.
Если сначала покрыты специальным сплетением гиф, покрывалом, - гемиангиокарпные. Два типа покрывала: общее и частное. Общее покрывает все плодовое тело (мухомор Amanita), частное соединяет края шляпки с ножкой (шампиньон Agaricus).
Гастероидные базидиомицеты: базидии созревают внутри плодовых тел и не отрываются активно от базидий. Они освобождаются в результате разрыва или общего разрушения плодового тела. Плодовые тела имеют хорошо выраженную оболочку перидий, одно, двуслойный, трехслойный. В трехслойном внешний слой – экзоперидий, средний – мезоперидий, внутренний – эндоперидий. Внутри перидия фертильная часть плодового тела – глеба. Может быть мономитической или димитической. В Глебе образуются полости – трамы. В камеры вдавливаются базидии с базидиоспорами. Типы развития и расположения спороносного слоя в глебе: 1)равномерный 2)локунарный 3)кораллоидный 4) многошляпочный 5) одношляпочный
Распространение спор за счет ветра, воды, насекомых, животных.
7) Циклы развития грибов: анаморфа и телеоморфа. Плеоморфизм. Варианты смены ядерных фаз: гаплофазный, диплофазный, гапло-диплофазный, гаплофазный с дикариофазой, гапло-дикариофазный и дикариофазный циклы развития. Анаморфный цикл развития. Понятие об анаморфных грибах.
Плеоморфизм – формирование в жизненном цикле нескольких спороношений разного облика. Половое спороношение (телеоморфа) обычно бывает только одно, а бесполых спороношений может быть одно или несколько.
Гаплофазный цикл (на примере Chytridiomycota, Synchytrium endobioticum – возбудителя рака картофеля): зооспора садится на поверхность молодого клубня картофеля и, не одеваясь оболочкой, переливается в одну из его эпидермальных клеток, которая разрастается (но не делится), а соседние усиленно размножаются. Паразит со временем одевается двухслойной оболочкой, превращаясь в летнюю цисту, иди просорус. Она образует вырост, в который переходят цитоплазма и ядро. Последнее сейчас же делится, а затем содержимое соруса распадается внутри общей оболочки на 5-7 многоядерных клеток. Число ядер там еще увеличивается, затем образуются зооспоры. Те же зооспоры из перезрелых сорусов могут попарно копулировать с немедленным слиянием ядер. Планозигота заражает хозяина и стимулирует деление его клеток в тангентальном направлении. Оказавшись таким образом в глубине, планозигота превращается в зимнюю, или покоящуюся, цисту, которая непосрдественно прорастает зооспорами (не образуя соруса).
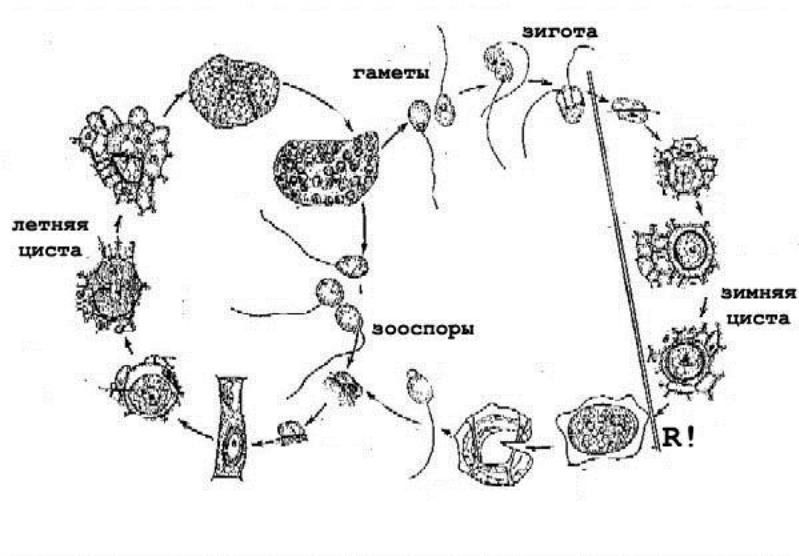
Гаплофазный цикл на примере Zygomycota, Mucor sp. Так называемая зигогамия: концы двух гиф сближаются (гифы разного знака, гетероталлизм), и на каждом конце отделяется по клетке, эти клетки сливаются между собой. Получившаяся зигота – зигоспора. Она прорастает в зародышевый спорангий (при прорастании происходит мейоз).
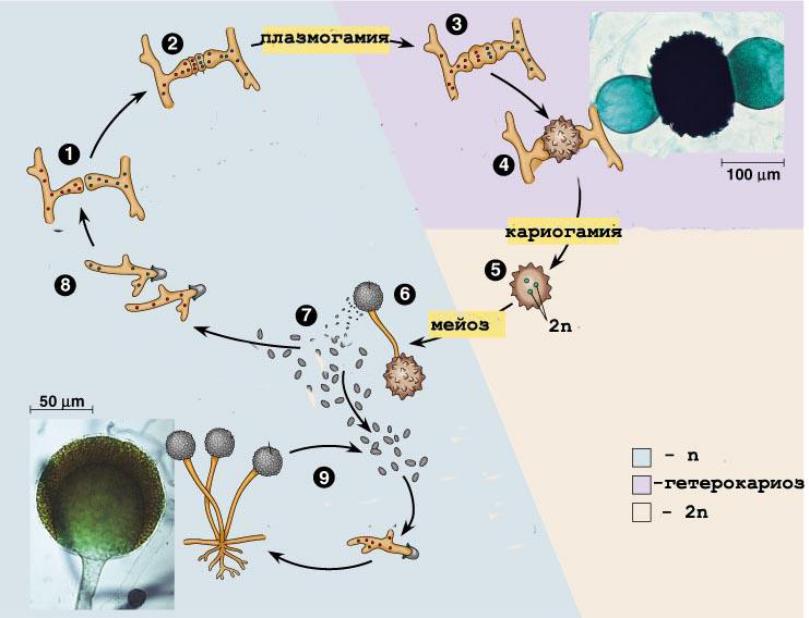
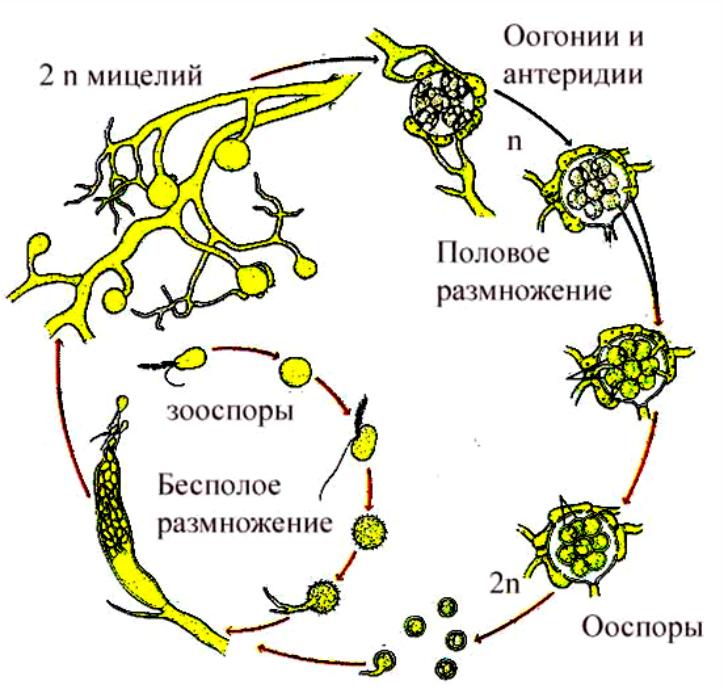
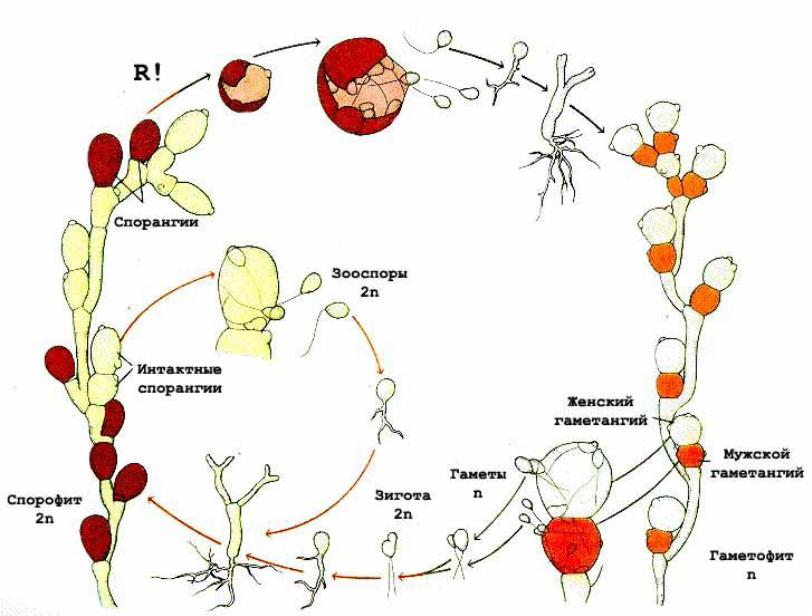
Диплофазный цикл на примере Oomycota, Saprolegina sp. Явление диморфизма. Из спорангия, развивающегося на конце гифы и отделяющегося от него перегородкой, выходят грушевидные зооспоры с двумя жгутиками на переднем конце. Поплавав некоторое время, они останавливаются и инцистируются (циста!), а затем одеваются оболочкой. Из оболочки выходит почковидная зооспора с двумя боковыми жгутиками. Она относительно долго плавает, затем садится на субстрат и прорастает в гифу. При половом процессии образуются оогонии (отделяющиеся перегородкой, одна или несколько яйцеклеток) и антеридии (на вершине специальных гиф – антеридиальных ветвей). Антеридиальные ветви подрастают к оогонию, антеридий плотно прижимается к нему и через поры в оболочке оогония пускает в него оплодотворяющие выросты. Развиваются ооспоры, одетые толстой оболочкой. После периода покоя прорастают в короткую гифу с зооспорангием на конце.
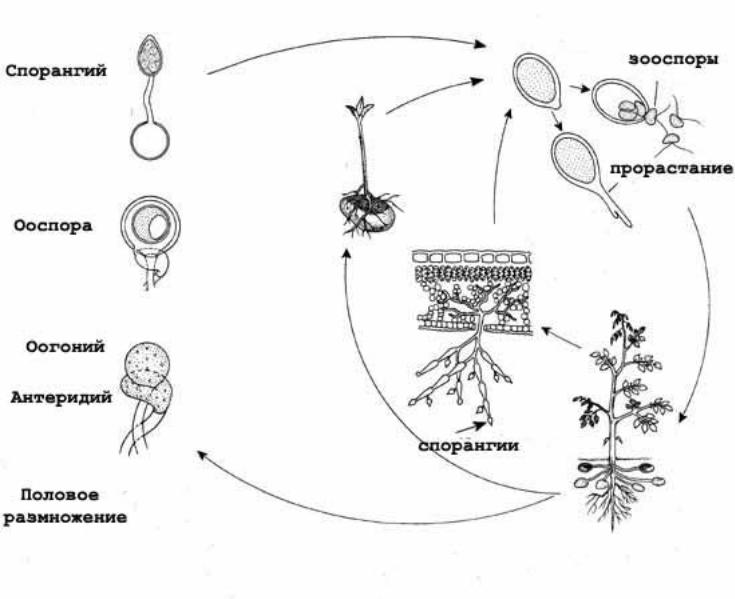
Диплофазный цикл на примере Oomycota: Phytophthora infestans.
Зооспорангии яйцевидные с сосочком на вершине, вскрывается крышечкой. Зооспорангии обычно опадают со спорангиеносцев и разносятся ветром и токами воды. При наличии капельно-жидкой воды зооспорангий прорастает зооспорами. При отсутствии – прорастает как экзогенная спора, конидия, непосредственно в гифу.
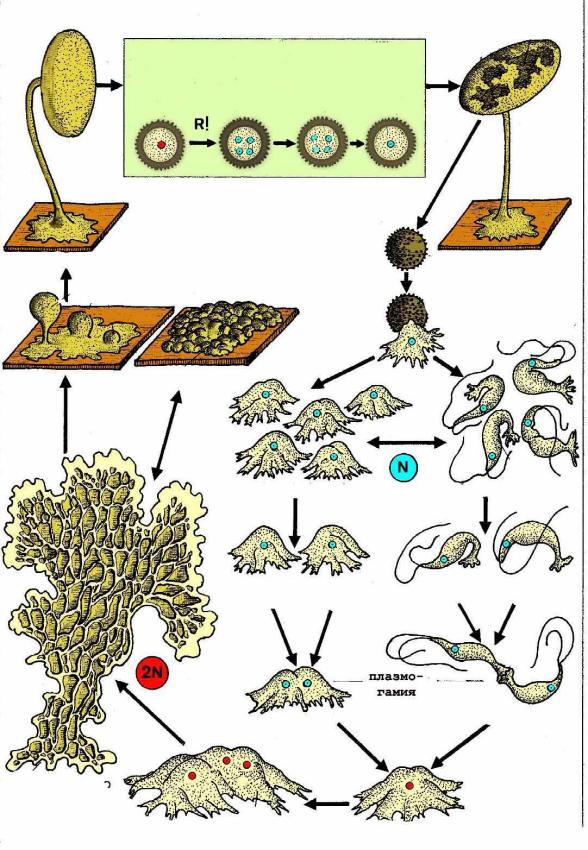
Гапло-диплофазный цикл на примере Myxomycota.
В определенный момент плазмодий (с диплоидными ядрами) приобретает положительный фототаксис, выползает на поверхность субстрата и переходит к генеративной стадии – образованию спорангиев со спорами (ядра редукционно делятся). Внутри спорангиев образуется масса спор и часто особые нити – капиллиции с неравномерно утолщенными стенками, способствуют разрыхлению и рассеиванию спор. Гаплоидные споры, попав на субстрат, прорастают в гаплоидные зооспоры с двумя гладкими жгутиками на переднем конце. Они могут делиться, затем теряют жгутики и превращаются в гаплоидные миксоамебы, которые сливаются попарно и образуются диплоидные миксоамебы. Она разрастается, число ядер увеличивается, вновь образуется плазмодий и уходит вглубь субстрата.
Гапло-диплофазный цикл на примере Chytridiomycota, Allomyces sp.
Изоморфная смена поколений. На диплоидном мицелии развиваются спорангии и покоящиеся клетки, которые тоже могут превращаться в зооспорангии. Если зооспорангий развивается из покоящихся клеток, то из него выходят гаплоидные зооспоры, развивающиеся в такой же по облику гаметофит, на котором имеются гаметангии, расположенные цепочками и разные по облику: одни с более крупными (женскими) гаметами, а другие, сидящие рядом с ними, - с гаметами меньшего размера (мужскими). Выйдя из гаметангиев, гаметы копулируют, и зигота без периода покоя сразу же прорастает в новую особь, минуя мейоз. Эта особь развивает только зооспорангии и покоящиеся клетки. Зооспоры, возникающие из настоящих зооспорангиев, похожи на гаметы, но крупнее их и диплоидны.
Гаплофазный цикл с дикариофазой
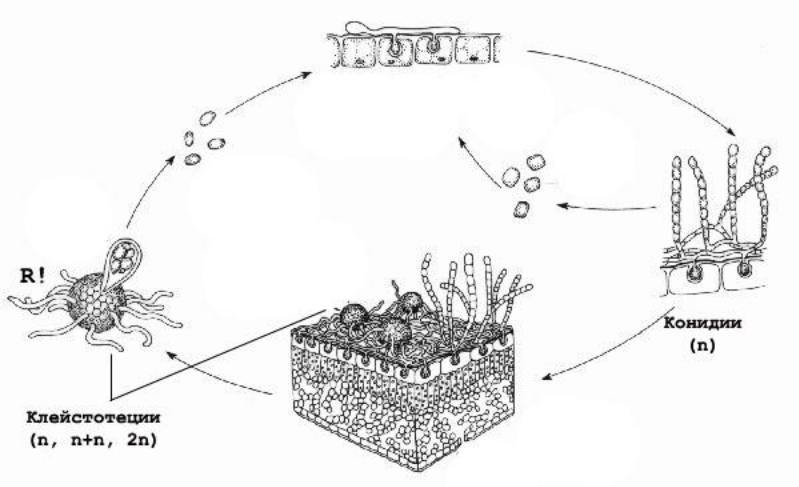
1) на примере Ascomycota, Erysiphales.
На мицелии на конидиеносцах развиваются конидии – в цепочках или поодиночке. Грают роль в возобновлении инфекции летом. «мучнистая роса»
Образование замкнутого плодовго тела – клейстотеция. Развитие начинается с образования половых органов в виде выростов на соседних гифах поверхностного мицелия. Архикарп не имеет трихогины и состоит из одного аскона, представляющего собой одноядерную клетку. Антеридий делится на ножку и собственно антеридий, который прикладывается к верхней части аскогона и переливает туда свое ядро. Происходит кариогамия, образуется 8 аскоспор.
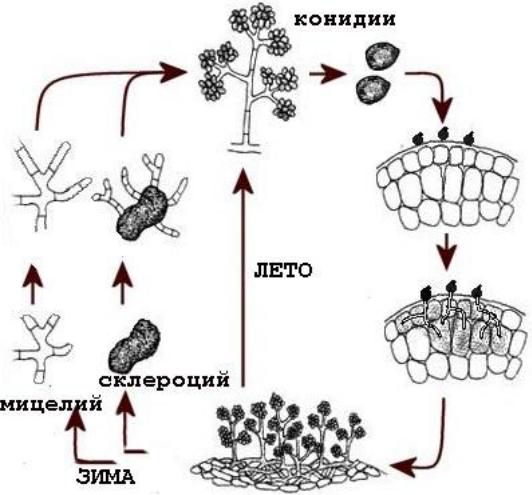
2) на примере Ascomycota, Claiceps purpurea. Спорынья.
Во время цветения заражаются аскоспорами цветки злаков. Мицелий проникает в нижнюю часть завязи, пронизывает ее своими гифами и вскоре образует конидиальное спороношение. Гифы сплетаются плотнее, образуя склероций черно-фиолетового цвета. Созревшие склероции падают на землю и зимуют. Весной они прорастают, образуя каждый 20-30 стром (ножка и головка). В головке формируются половые органы →копуляция→перитеций.
3) на примере Ascomycota, Ventutia inaequales
Парша яблони. Сумчатые спороношения развиваются только на опавших листьях после зимовки. При этом мицелий проникает в толщу листа, образуются клубочки – зачатки псевдотециев. Внутри клубочка дифференцируется архикарп в виде закрученной толстой гифы, ее конец выходит наружу и вытягивается в трихогину. Рядом развивается антеридий, верхняя многоядерная клетка которого дает лопасти и охватывает ими верхнюю часть трихогины, мужчские ядра переходят в трихогину.
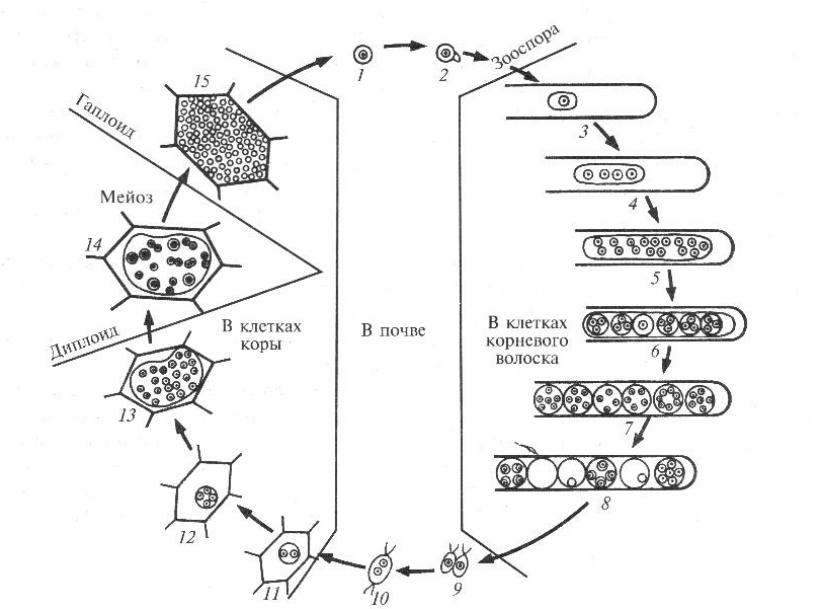
Гапло-дикариофазный цикл на примере Basidiomycota: Uredinales, Puccinia graminis
Весной телиоспоры прорастают в фрагмобазидию с четыремя базидиоспорами разных половых знаков. Перед прорастанием дикариотичные ядра сливаются, и диплоидное ядро делится редукционно. Гаплоидные ядра проходят в базидиспоры. Те сбрасываются и попадают в воздушные потоки. Попадают на листья барбариса. На верхней поверхности листа базидиоспоры прорастают в гаплоидный мицелий, образуется спермогонии (пикни), при созревании там образуются споры спермации или пикниспоры. Из спермогония наружу выступают гифы перифизы, к ним прилипают спермации. Насекомые переносят их с одно пикни на другую. В итоге внутри тканей барбариса развиваются дикариотические гифы, образующие клубочки внутри тканей листьев, которые затем формируют эции с нижней стороны листьев. Образуются эциоспоры гриба. Переносятся на злаки воздушными потоками. Там образуются урединии с урединоспорами. Которые снова и снова заражают злаки. А в конце лета здесь же формируются телиоспоры…
Дикариофазный цикл на примере Basidiomycota: Ustilaginales
Заражение происходит в дикариофазе! Устоспоры, содержащие по 2 ядра дикариона, попадают в почву с посевными зернами. Прорастают в базидию, мейоз, гаплоидные базидиоспоры. Копулируют между собой, далее развивается дикариотичный мицелий. Он заражает растение, продвигается вверх, поражая узлы и междоузлия растения. Далее в определенных местах образует устоспоры.

Анаморфный цикл развития. Анаморфные грибы.
Размножаются только бесполым путем – конидиями или вегетативно – почкованием. Нет половой стадии. Эволюция направлена на усовершенствование конидиального аппарата. Отдел Deutoromycota, 3 класса.
8) Экология грибов. Влияние основных экологических факторов на развитие грибов. Питание грибов. Трофические группы (биотрофы: паразиты и симбиотрофы; некротрофы, сапротрофы) Паразитизм (адаптации грибов к паразитическому образу жизни, облигатный и факультативный паразитизм). Микориза и ее значение в формировании природных сообществ. Субстратные группы грибов (ксилотрофы, подстилочные сапротрофы, копротрофы, энтомофилы, микофилы и т.д.). Представление о сообществах почвенных и водных грибов.
Экологические группы грибов характеризуют их распределение по субстрату, который является источником питания грибов. Критерии выделения конкретных групп могут быть предметом дискуссий.
В микологической литературе выделяют следующие группы макромицетов:
симбиотрофные макромицеты (микоризообразователи) — макромицеты, образующие микоризу на корнях деревьев и кустарников.
Микориза, грибокорень, симбиоз мицелия гриба и корней высшего растения. М. могут образовывать некоторые зигомицеты, аскомицеты (трюфелевые, Тuberales) и гл. обр. базидиальные грибы (агариковые и болетовые). Различают М. эктотрофную, при которой гриб оплетает корень, оставаясь на его поверхности (М. базидиальных грибов — болет, сыроежка, паутинник и др.— с лесными деревьями) и эндотрофную, когда гриб проникает внутрь корня (М. микроскопич. грибов из класса несовершенных с растениями сем. орхидных и вересковых). Тифы гриба в клетках древовидно ветвятся или образуют головчатые вздутия. М. рассматривают либо как мутуалистический симбиоз, от котоpoгo выгоду получает и гриб и растение, либо как ограниченный паразитизм. Грибы-микоризообразователи, вероятно, разлагают некоторые недоступные растению органич. соединения почвы, способствуют усвоению фосфатов, соединений азота, вырабатывают вещества типа витаминов и активаторы роста, а сами используют вещества (возможно, углеводы), извлекаемые ими из корня растения. Семена нек-рых растений (напр., орхидных) способны прорастать только в присутствии гриба.
сапротрофные макромицеты (сапротрофы) — макромицеты, использующие в качестве источника пищи мёртвое органическое вещество, за счёт которого осуществляются все их процессы жизнедеятельности.
подстилочные и гумусовые сапротрофы — грибы, использующие для питания лесной опад, подстилку и гумусовый слой почвы.
ксилотрофы (дереворазрушающие грибы) — грибы, осуществляющие разложение древесины.
карботрофы — грибы, растущие на кострищах и пожарищах.
копротрофы — грибы, использующие для жизнедеятельности органику из экскрементов животных.
бриотрофы — грибы, разлагающие отмершие части мхов (если мхи сфагновые, то грибы имеют название сфагнотрофы).
микотрофы (сапротрофные микофилы) — грибы, развивающиеся на мумифицированных плодовых телах шляпочных грибов (в основном, груздях и сыроежках).
Грибы очень специфичны по своим требованиям к условиям роста и развития. Они обычно строго приурочены к комплексу экологических условий (особенно к субстрату), в которых данный род или вид имеет преимущества перед другими родами или видами. Грибы питаются готовыми органическими веществами, поэтому экологические группы грибов выделяются по их приуроченности к тому или иному субстрату — одному из основных экологических факторов для гетеротрофных организмов. По способу питания, как уже говорилось, различают грибы-паразиты, питающиеся органическими веществами живых организмов (растений и животных), и грибы-сапротрофы, живущие за счет мертвого органического материала.
Питание
Все грибы являются гетеротрофными организмами. Минеральные вещества гриб способен усваивать из окружающей среды, однако органические он должен получать в готовом виде. В зависимости от потребности в веществах, тот или иной вид грибов заселяет определённый субстрат. Грибы не способны усваивать крупные частички пищи, поэтому всасывают исключительно жидкие вещества через всю поверхность тела, при этом огромная площадь поверхности мицелия оказывается весьма выгодной.
Внешнее пищеварение у грибов
Для грибов характерно внешнее пищеварение, то есть сначала в окружающую среду, содержащую пищевые вещества, выделяются ферменты, которые вне организма расщепляют полимеры до легкоусвояемых мономеров, которые всасываются в цитоплазму.
9) Лихенизированные грибы (лишайники). Особенности морфологии (накипные, листоватые и кустистые слоевища). Анатомическое строение слоевища (гомеомерное и гетеромерное). Особенности микобионта и фотобионта. Размножение лишайников (изидии, соредии, строение плодовых тел). Биохимические особенности (лишайниковые вещества). Взаимоотношения компонентов. Распространение лишайников. Субстратные группы лишайников.
Общая характеристика. Лишайники - это своеобразная группа живых организмов, тело (слоевище) которых образовано двумя организмами — грибом (микобионт) (класс и водорослью или цианобактерией (фикобионт), находящимися в симбиозе. В составе лишайников обнаружено около 20 тыс. видов грибов и около 26 родов фототрофных организмов. Наиболее часто встречаются зеленые водоросли родов требуксия, трентеполия и цианобактерия носток, являющиеся авто-трофными компонентами примерно у 90% всех видов лишайников.
Симбиотические (мутуалистические) взаимоотношения между компонентами лишайников сводятся к тому, что фикобионт снабжает гриб созданными им в процессе фотосинтеза органическими веществами, а получает от него воду с растворенными минеральными солями. Кроме того, гриб защищает фикобионт от высыхания. Такая комплексная природа лишайников позволяет им получать питание из воздуха, атмосферных осадков, влага росы и туманов, частиц пыли, оседающей на слоевище, из почвы. Поэтому лишайники обладают уникальной способностью существовать в крайне неблагоприятных условиях, часто совершенно непригодных для других организмов, — на голых скалах и камнях, крышах домов, заборах, коре деревьев и др.
Микобионт отличается специфичностью, т. е. входит в состав только одного вида лишайника.
Строение лишайников. Слоевище лишайников обычно серого, светлого или темно-бурого цвета. По внешнему виду талломы лишайников делятся на накипные, листоватые и кустистые .
Наиболее распространены накипные, или корковые, лишайники (около 80%), имеющие таллом в виде тонкой корочки, прочно срастающейся с субстратом и не отделимой от него. Более высокоорганизованные листоватые лишайники имеют вид чешуек или пластинок, прикрепляющихся к субстрату пучками гиф, называемых ризинами. Они разрастаются на камнях и коре деревьев. Так, например, на стволах и ветках осины часто встречается лишайник золотистого цвета—ксантория. Кустистые лишайники представляют собой кустики, образованные тонкими ветвящимися нитями или стволиками, прикрепленными к субстрату лишь основанием.
По анатомическому строению лишайники делят на гомео- и гетеромерные. У гомеомерных лишайников слоевище представляет собой рыхлое сплетение гиф гриба, среди которых более или менее равномерно располагаются клетки или нити фикобионта.
В гетеромерном слоевище четко различимы слои: кора, слой фотобионта и сердцевина. Гетеромерное строение характеризуется наличием в слоевище дифференцированных слоев, каждый из которых выполняет определенную функцию: верхняя и нижняя кора — защитную, фотосинтезирующий слой участвует в процессе фотосинтеза и накапливает продукты ассимиляции. Такой морфологический тип лишайника представляет собой наиболее высокоорганизованную форму слоевища и характерен для большинства листоватых и кустистых лишайников.
Микобионты лишайников продуцируют вторичные метаболиты – лишайниковые кислоты.
В апотециях и перитециях формируются споры для размножения лишайникового гриба.
Перитеции встречаются у небольшого числа лишайников. Апотеции образуются у многих видов лишайников. Они имеют чаще блюдцевидную форму, но встречаются и более или менее выпуклые или почти шаровидные. Самые крупные апотеции имеют диаметр более 1 см, но лишайников с такими апотециями немного. У громадного большинства лишайников диаметр апотециев от одного до нескольких миллиметров. Ониразбросаны по поверхности листоватого таллома, чаще в его середине, или расположены по краям его лопастей. У кустистых лишайников апотеции обычно находятся на концах веточек или сциф. Они бывают сидячими или реже на небольших ножках приподнимаются над талломом. Апотеции могут быть окрашены в один цвет с талломом, или поверхность апотеция (так называемый диск апотеция) имеет другую окраску.
По строению различают три типа апотециев: леканориновый, лецидеиновый и биаториновый.
Леканориновый апотеций по своему анатомическому строению сходен с талломом лишайника. Его диск имеет талломный (слоевищный) край, образованный талломом лишайника и состоящий из гиф гриба и клеток водоросли. Этот край окрашен так же, как и таллом лишайника, и отличается по окраске от самого диска. В самом талломе под леканориновым апотецием также имеются клетки водоросли.
Лецидеиновый апотеции имеет край, состоящий только из гиф гриба и окрашенный в тот же цвет, что и диск апотеция. В этом крае, а также в талломе под лецидеиновым апотецием клеток водорослей нет. Лецидеиновые апотеции имеют твердую консистенцию и обычно темную окраску.
Биаториновый апотеций имеет также же строение, как лецидеиновый, но отличается яркой окраской и мягкой консистенцией.
Вегетативное размножение:
Соредии – мелкие комочки, состоящие из одной или нескольких клеток фотобионта, окруженных гифами гриба.
Изидии – выросты слоевища, состоящие как из гиф гриба, так и из клеток фотобионта.
В целом лишайники неприхотливы. Лимитирующим фактором является загрязнении окружающей среды. Экологические группы:
- эпигейные лишайники – обитают на почвах, бедных питательными веществами (например, песчаных). В основном кустистые.
- эпифитные – на коре стволов, ветвей деревьев и кустарников. Листоватые, накипные, куститстые.
- эпифильные – на листьях вечнозеленых пород.
- эпиксильные – на древесине мертвых деревьев
- эпилитные – на камнях
- водные лишайники
10) Основы систематики грибов. Традиционные и современные подходы к классификации. Геносистематика и ее значение для микологии. Основные взгляды на происхождение и пути эволюции грибов и грибоподобных организмов. Роль грибов в эволюции высших растений.
Новая систематика
Грибы
|
Uniconta |
|||
|
Opistoconta |
|||
Царство |
Fungi |
|||
Отдел |
Chytridiomycota |
Zygomycota |
Ascomycota |
Basidiomycota |
Класс |
Chytridiomycetes |
Zygomycetes |
|
|
Порядок |
Chytridiales |
Mucorales |
|
|
Представители |
Synchitrium endobioticum |
Mucor, Zyzygites |
|
|
Отдел Ascomycota
Подотделы |
Taphrinomycotina |
Saccharomycotina |
Pezizomycotina |
Класс |
Taphrinomycetes |
Saccharomycetes |
|
Порядок |
Taphrinales |
Saccharomycetales |
|
Представители |
Taphrina padi |
Saccharomyces cerevisiae |
|
Подотдел Pezizomycotina
Класс |
Eurotiomycetes |
Erysiphomycetes |
Sordariomycetes |
Peziziomycetes |
Leotiomycetes |
Dothideomycetes |
Порядок |
Eurotiales |
Erysiphales |
|
Pezizales |
|
Pleosporales |
Представители |
Eupenicillium |
Microsphaera palzchewskii Uncinula adunka |
|
Peziza Morchella Gyromitra Tuber |
|
Pleospora Venturia inaequales |
Класс Sordariomycetes
Порядок |
Sordariales |
Xylariales |
Hypocreales |
Представители |
Sordaria |
Xylaria Hypoxylon |
Claviceps purpurea Epichloe typhina |
Класс Leotiomycetes
Порядок |
Helotiales |
Rhytismatales |
Представители |
Monilinia fructigena |
Lophodermium |
Отдел Basidiomycota
Класс |
Basidiomycetes П/кл Homobasidiomycetidae |
Urediniomycetes |
Ustilaginomycetes |
|
Порядок |
|
Uredinales |
Ustilaginales |
Tilletiales |
Представители |
|
Puccinia graminis Triphragmium ulmariae Phragmidium rubi-idaei |
Ustilago tritici |
Tilletia |
Класс |
Basidiomycetes |
|||
Подкласс |
Homobasidiomycetidae |
|||
Порядок |
Polyporales |
Clavariales |
Cantharellales |
Lycoperdales |
Представители |
Fomes fomentarius Daedalia quercina |
Clavaria |
Hydnum Hydnellum |
Lycoperdon Bovista Geastrum Crucibulum |
Грибоподобные организмы
|
Uniconta |
|
Царство |
Amoebozoa |
|
Отдел |
Myxomycota |
|
Класс |
Myxomycetes |
|
Порядок |
Liciales |
Stemonitales |
Представители |
Lycogala |
Stemonites |
|
Rhizaria |
Царство |
Cercozoa |
Отдел |
Plasmodiophoromycota |
Класс |
Plasmodiophoromycetes |
Порядок |
Plasmodiophoromycetales |
Представитель |
Plasmodiophora brassicae |
Царство |
Stramenopila |
|
Отдел |
Oomycota |
|
Класс |
Oomycetes |
|
Порядок |
Saprolegniales |
Peronosporales |
Представители |
Saprolegnia |
Peronospora Phytophtora infestans Plasmopara aegopodii |
11) Amoebozoa
Отдел Myxomycota (Слизевики) – положение в системе, особенности строения и размножения. Основные представители и их циклы развития.
Царство Amoebozoa (Protozoa)
Отдел Myxomycota
Класс Myxomycetes
Порядок Liciales: Lycogala sp.
Порядок Stemonitales: Stemonites sp.
Порядок Physarales
Общая характеристика:
В вегетативном состоянии – амебоидные организмы, плазмодий.
При размножении плазмодий превращается в различного рода вместилища, содержащие массу спор. При прорастании споры выходящий из нее голый протопласт может быть либо одноядерной амебоидной клеткой (миксоамебой), либо представлять собой зооспору (планоцит), снабженную одним или двумя передними жгутиками. Обе формы способны к размножению делением. Сливаются попарно со слиянием ядер, образуя зиготу. Зигота живет как голая амебоидная клетка, поглощает пищу и увеличивается в размере. Митотические деления ядра, получается плазмодий.
В вегетативном состоянии их представители диплоидны, только перед образованием спор происходит мейоз, поэтому инцистированные споры содержат гаплоидные ядра.
Обитают на почве и мертвом орг. субстрате и питаются путем внутреннего переваривания частиц пищи.
Репродуктивная стадия – простой или сложный спорангий.
В составе клеточных стенок спорангиев и спор есть целлюлоза.
Класс миксомсицеты Myxomycotes
Вегетативное тело – многоядерный плазмодий. Он передвигается от света (отрицательный фототаксис) в сырые места (положительный гидротаксис), скрыт в толще субстрата. Актин участвует в движении (0,1 мм в мин). 3 типа плазмодиев:
- протоплазмодий. Микроскопические размеры, нет токов цитоплазмы.
- афаноплазмодий. Отсутствие слизистого чехла, прозрачен, менее устойчив к обезвоживанию.
- фанероплазмодий. Более плотный, вееровидной формы, сетчатая структура, ток цитоплазмы, защитный слизистый чехол, часто окрашен.
Гапло-диплофазный цикл на примере Myxomycotetes.
В определенный момент плазмодий (с диплоидными ядрами) приобретает положительный фототаксис, выползает на поверхность субстрата и переходит к генеративной стадии – образованию спорангиев со спорами (ядра редукционно делятся). Внутри спорангиев образуется масса спор и часто особые нити – капиллиции с неравномерно утолщенными стенками, способствуют разрыхлению и рассеиванию спор (сплошные или полые трубки капиллиция, анастамозы, развивается из вакуолей, важен в систематике). Спорангии образуются обыкновенно группами на пов-ти плазможия и потребляют все его содержимое. Если срастаются совсем плотно, то образуются массивыне округлые тела – эталии, окруженные общей оболочкой. Внутри – псевдокапиллиций, который увеличивает прочность спороношения. Гаплоидные споры, попав на субстрат, прорастают в гаплоидные зооспоры с двумя гладкими жгутиками на переднем конце. Они могут делиться, затем теряют жгутики и превращаются в гаплоидные миксоамебы, которые сливаются попарно и образуются диплоидные миксоамебы. Она разрастается, число ядер увеличивается, вновь образуется плазмодий и уходит вглубь субстрата.
Споры – микроскопически малые шаровидные клетки, одетые твердой оболочкой.
Порядок физаровые – Physarales.
Самый большой по числу видов слизевиков. Споры темные, плодоношения – спорангии, сросшиеся спорангии, эталии. Имеют известковые гранулы в спорангиях и в капиллиции. Ярко окрашенные плазмодии.
Порядок стемонитовые - Stemonitales
Плазмодий очень нежный, прозрачный. Спороношение – отдельные спорангии или эталии. Капиллиции – сетчато анастомозирующие нити. Спорнагии сидят на ножке + колонка, от которой отходят капиллиции. Споры темные. Stemobites sp – встречается на гнилой древесине, засохших листьях и т.д.
Порядок Liceales. Лицевые.
Спорангии, эталии, плазмодиокарпы. Споры светлые. Нет настоящего капиллиция – только псевдо (система ветвящихся трубок). Lycogala sp. «волчье вымя» на сильно сгнивших пнях, стволах. Плазмодий кораллово-красный. Эталии группами. Молодые эталии нежно-розовые, далее темнеют, сверху образуется отверстие, и вылетают споры.
Порядок Trichiales трихиевые: яркая окраска, меняется в процессе созревания, спорангии одиночные.
Происхождение: скорее всего от древних амеб.
Дикариотический(гапло-диплофазный цикл). У грибов встречается редко и описана у некоторых водных хитридиомицетов и псевдогрибов. Рассмотрим данный цикл на примере myxomycota. Основной вариант существования – плазмодий. Стоит упомянуть, что ядра в плазмодии всегда диплоидные. В случае неблагоприятных условий плазмодий может переходит в стадию покоя, покрываясь корочкой. При переходе к размножению образуется спорангий. За закрытыми дверями происходит меойз: формирование 4 гаплоидных ядер. Жизнь несправедлива – 3 из них погибают. Споры гаплоидные в конце. Из них выходят маленькие гаплоидные жгутиковые стадии(зооспора=планоцит)\амебоиды(миксамёбы). Функция данной стадии – исключительно расселительная. Спустя недолгое время начнут выполнять функцию гамет. Сперва происходит плазмогамия и практически сразу кариогамия. Образуется диплоидный амебоид. Далее следует нечто удивительное – слияние в многоядерный трофоплазмодий. Если рассматривать как основной вариант строения, то нельзя четко сказать – гаплоидная или диплоидная стадия.
12) CERCOZOA
Отдел Plasmodiophoromycota – паразитические слизевики. Основные представители и их циклы развития.
Царство Cercosoa
Отдел Plasmodiophoramycota
Класс Plasmodiophoromycetes
Порядок Plasmodiophoromycerales
Plasmodiophora brassicae


На корне капусты образуются опухоли. Темные клетки с зернистым содержимым. Некоторые клетки полностью заполняются спорами из плазмодия. Корень сгнивает. На след. год споры прорастают зооспорами, которые прикрепляются к корневым волоскам капусты. Зооспоры инцистируются. Внутри образуется трубка, которая прикрепляется к стенке клетки хозяина, пронзает ее. Проникновение внутрь хозяина. Там возникает миксоамеба, превращается в гаплоидный плазмодий с многими ядрами. Он делится и распространяется из клетки в клетку. Позже распадается на одноядерные части, которые делятся и становятся вторичными зооспорами ли гаметами. Могут заражать хозяина непосредственно (n) или после копуляции (2n).
Для борьбы с паразитом меняют реакцию почвы с кислой (благоприятной для них) на щелочную.


13) Stramenopila
Отдел Oomycota. Взгляды на происхождение и положение в системе. Строение мицелия и состав клеточной стенки. Размножение (бесполое: двужгутиковые зооспоры и конидии, и половое: оогамия). Особенности эволюции в связи с выходом на сушу и переходом к паразитизму. Диплофазный цикл развития.
Порядок Saprolegniales. Основные представители и особенности жизненных циклов (дипланетизм, тенденция к утрате жгутиковых стадий как одно из направлений эволюции).
Порядок Peronosporales. Адаптации к паразитическому образу жизни. Переход к конидиальному спороношению и его биологический смысл. Основные представители, их циклы развитияи, практическое значение.
Царство Straminopila
Отдел Oomycota
Порядок Saprolegniales: Saprolegnia sp
Порядок Peronosporales: Phytophtora sp, Plasmopara, Peronospora.
Характеристика
1. Вегетативное тело – хорошо развитый несептированный мицелий, иногда ризомицелий.
2. Бесполое размножение посредством зооспор с двумя или одним передним жгутиком. У наиболее прогрессивных форм зооспорангий отрывается от мицелия целиком и прорастает в гифу.
3. Половой процесс – изогамия или особенная оогамия, при которой антеридий не дифференцируется на гаметы.
4. Таксон включает около 700 видов. Сапротрофы и паразиты. Прослеживается выработка приспособлений, связанная с переходом на наземный образ жизни.
Класс Oomycetes.
Хорошо развит несептированный мицелий, отдельные фрагменты легко регенерируют. Бесполое размножение посредством спор, у Peronosporales зооспорангий, не дифференцируясь, прорастает в гифу, т.е. функционирует как конидия. Половой процесс – особенная оогамия. В основном водные обитатели, но есть и наземной среды. Сапротрофы и паразиты (переход к наземному обитанию).
Порядок Сапролегниевые. Saprolegniales.
100 видов. Вегетативное тело состоит из толстых свободных гиф и ветвящихся тонких ризоидов, прикрепляющихся к субстрату. В кл. стенке целлюлоза. На концах свободных гиф развиваются зооспорангии, антеридии, оогонии. При неблагоприятных условиях гифы образуют геммы – особые хламидоспоры, с густым содержимым и толстыми оболочками. В оогониях несколько яйцеклеток, на их образование идет все содержимаое оогоний. Антеридии – тонкие цилиндрические клетки. Преимущественно водные сапротрофы. Некоторые паразитируют на рыбах и лягушках.
Характерно явление диморфизма (способность менять положение жгутиков) и дипланетизм (способность зооспор инцистироваться, а потом снова становится активными).
Диплоидный цикл – вегетативный таллом диплоидный, меойз – при формировании гаметангиев или гамет(гаметический). Характерен для оомицетов и некоторых сумчатых дрожжей.
Рассмотрим данный процесс на примере отдела Oomycota: Saprolegnia sp. Анаморфный вериант размножения(эукарпический тип образования зооспор) – образование и выход из мицелия грушевидной зооспоры, обладающей гетероконтными жгутиками(передний конец). Утрата жгутиков после непродолжительного плавания и образования «лжестадии отдыха». Уже после непродолжительного отдыха выходит зооспора бобовидной формы, обладающая опять же гетерокотными жгутиками на боковой поверхности. Она то и расселяется и прорастает без редукционного деления. На том же самом диплоидном мицелии образуется и половые органы – гаметангии. Оогонии(женские) и антеридии(мужские). Данные половые органы – многоядерные, дифференцированные, гаплоидные(в результате мейоза= гаметическая редукция). Антеридий подрастает к оогонии и выпускает свое внутренне содержимое(равное количеству яйцеклеток внутри оогонии). Происходит сперма плазмогамия, но и кариогамия не долго заставляет себя ждать. После оплодотворения образуются диплоидные ооспоры, одетые толстой оболочкой. Обычно такие ооспоры переходят в состояние покоя с могут переносить неблагоприятные условия. Ооспоры после периода покоя прорастают в короткую гифу с зооспорангиями на конце.
Порядок Peronosporales пероноспоровые.
Характерна эволюция бесполых спороношений от типично спорангиальных к конидиальным, что тесно связано с переходом от водного к наземному образу жизни. И переход от сапротрофного образа жизни к паразитическому. Эволюция: Зооспорангий, из которого выходят зооспоры с двумя жгутиками, →отрыв зооспорангия целиком с прорастанием в зооспоры→отрыв зооспорангия с прорастанием в гифу→зооспорангий не функционирует, просто прорастает в гифу (конидия).
Phytophtora infestans : спорангиеносцы пучками высовываются из устьица на нижней стороне листа. Паразит, возбудитель витофтоза пасленовых, например картофеля. «картофельный гриб». Мицелий идет по межклетникам, в клетки внедряются гаустории. На листьях появляются бурые пятна отмершей ткани. С нижний стороны листа по краю пятна заметен пушок – сплетения спорангиеносцев. Они несут зооспорангии, те отваливаются целиком и переносятся на новый лист или клубни.
Немного сложнее выглядит ситуация диплоидного цикла с представителями Ооmycota: Phytophthora infestans. Бесполое размножение здесь происходит несколькими способами. Зооспорангии обычно опадают с спорангионосцев и распространяются ветром или током воды. Зооспорангии лимоновидной формы с небольшим бугорком на вершине. При наличии капельно-жидкой среды зооспорангий прорастает сразу зооспорами. Но если погода нелетная и отсутствует даже стаканчик чистой воды – зооспорангий прорастает как отдельная экзогенная спора – конидия – непосредственно в гифу. Поэтому спорангиоспоры и спорангии пероноспоровых называют, соответственно конидиеносцами
и конидиями. При переходе к половому размножению на мицелии образуются антеридии и оогонии.Оба гаплоидные. Антеридий удивительным способом подрастает к оогонии и происходит оплодотворение. Ооспора образуется в данном случае. И при выходе в высший свет – прорастает спорангием. Который тоже, на удивление, дает зооспорангий.
Plasmopara : паразит на виноградной лозе, на подсолнечнике.
Peronospora : конидии, прорастают гифами. Поражают табак.
13) Mycota (Fungi)
Pазличные взгляды на происхождение группы и ее объем.
Отдел Chytridiomycota. Класс Chytridiomycetes. Особенности строения и размножения. Ультраструктурные особенности зооспор. Разнообразие форм полового процесса (хологамия, изогамия, гетерогамия и оогамия). Циклы развития (гаплофазный и гапло-диплофазный).
Порядок Chytridiales. Основные представители, их циклы развития и экологические особенности. Практическое значение.
Порядки Blastocladiales и Monoblepharidales. Особенности строения и размножения. Взгляды на положение в системе.
К настоящим грибам относятся хитридиомикота, зигомикота, аскомикота, базидиомикота.
Царство Mycota (Fungi)
Отдел Chytridiomycota
Класс Chytridiomycetes
Порядок Chytridiales : Synchytrium, Chytridium, Rhizophydium
Порядок Blastocladiales : Allomyces
Порядок Monoblepharidales :
Общая хар-ка
1. Примитивные представители не имеют мицелия – одноклеточные организмы. Другие имеют ризомицелий или несептированный мицелий.
2. Кл.стенка хитиново-глюкановая.
3. В цикле развития имеются жгутиковые стадии – зооспоры и гаметы с одним гладким жгутиком!
4. Бесполое размножение – зооспорами.
5. Половое: изогамия, гетерогамия, оогамия, слияние гаметангиев, соматогамия.
6. Гаплонты.
7. Преимущественно водные организмы. Паразиты на водорослях и низших животных или сапротрофы на субстратах.
8. около 1 000 видов.
Ультраструктура зооспор:
обычно 1 жгутик. Ядро одно. Его форма и расположение различны и могут служить диагностическим признаком. Возле кинетосомы жгутика лежит второе базальное тело – покоящееся. От кинетосомы вглубь цитоплазмы отходят МТ, образующие жгутиковые корни и ризопласты (более массивные). У представителей Monoblepharidales имеется специфическая органелла – румпосома, участвующая в работе жгутика.
Порядок Chytridiales хитридиевые
Свыше 700 видов. Либо одна клетка, либо ризомицелий. Сапротрофы живут на орг. остатках, содержащих хитин и кератин. Род Synchytrium – цисты прорастают в сорус спорангиев, образующийся из выроста первоначального вегетативного тела. Sunchytrium endobioticum – возбудитель рака картофеля (бугристые опухоли на клубнях).
Гаплоидный (гаплофазный) цикл – ядра в вегетативном талломе гаплоидны. Диплоидная зигота(обычно после периода покоя) делиться мейотически(зиготическая редукция) – зигомицеты, многие хитридиомицеты.
Описание цикла на примере Chytridiomycota: Synchytrium endobioticum: гаплоидные зооспоры со жгутиком(1,задний, гладкий) заражают картофель, попав в тело хозяина. Внутри полости тела хозяина образуется вегетативное тело, типа амебоид(1 гаплоидное ядро). Далее следует интенсивное деление n-ядер с образованием довольно крупного плазмодия. По мере истощения хозяина , плазмодий формирует плотную оболочку и переходит в состояние гаплоидной «летней цисты». После периода покоя формирует целую группу зооспорангиев с большим количеством зооспор. Эта стадия носит название сорус зооспор. Ближе к осени вышедшие зооспоры функционируют как гаметы. После слияние «гамет» образуется планозигота(подвижная) – дикариотическая. То есть наблюдаем сперва плазмогамию. После будет происходить кариогамия. Начинается непродолжительная диплоидная стадия – стадия «зимовки зимней цисты». После определенного периода покоя происходит редукционное деление(мейоз и последующая очередь митозов) – и образование гаплоидных зооспор, готовых далее захватывать мир!
Порядок Blastocladiales : сапротрофы на растительных остатках и трупах насекомых в воде. В талломе различимы главная ветвь и боковые ветви. Хаар-но образование толстостенных покоящихся спорангиев с ямчатой поверхностью. Подвижные стадии – зооспоры и гаметы. Allomyces sp. Изоморфная смена поколений.
Гапло-диплофазный цикл на примере Chytridiomycota, Allomyces sp.
Изоморфная смена поколений. На диплоидном мицелии развиваются спорангии и покоящиеся клетки, которые тоже могут превращаться в зооспорангии. Если зооспорангий развивается из покоящихся клеток, то из него выходят гаплоидные зооспоры, развивающиеся в такой же по облику гаметофит, на котором имеются гаметангии, расположенные цепочками и разные по облику: одни с более крупными (женскими) гаметами, а другие, сидящие рядом с ними, - с гаметами меньшего размера (мужскими). Выйдя из гаметангиев, гаметы копулируют, и зигота без периода покоя сразу же прорастает в новую особь, минуя мейоз. Эта особь развивает только зооспорангии и покоящиеся клетки. Зооспоры, возникающие из настоящих зооспорангиев, похожи на гаметы, но крупнее их и диплоидны.
Если наш мицелий диплоидный, то это спорофит. Наличие 2 «типов» спорангий. Одни из спорангиев – интактные – образуются интактные диплоидные зооспоры, которые прорастают точно таким же мицелием! Так же на этом же мицелии присутствуют другие спорангии. В которых после спорической редукции образуются гаплоидные споры, дающие начало гаметофитам. Наличие мужской и женской гаметангии на одном гаметофите. Формируются гаметы 2 тмпов – подвижные, но разные по форме. После недолгих поисков происходит слияние гатем с образованием планозиготы, которая оседает и прорастает в спорофит. 2 отростка прорастающей планозиготы – закладывается дифференциация.
Порядок Monoblepharidales: многоядерный мицелий без перегородок, состоящий из ризоидов и свободных длинных гиф. В кл.стенках нет хитина. Органы бесполого размножения – зооспорангии. Настоящая оогамия. Половые органы – оогонии (где развивается одна или несколько яйцеклеток) и антеридии (4-8 сперматозоидов). Располагаются на одной и той же гифе рядом друг с другом. Зигота после периода покоя прорастает в новую гифу. Гаплонты. Пресноводные сапротрофы. Имеется специфическая органелла – румпосома, участвующая в работе жгутика.
14) Отдел Zygomycota. Класс Zygomycetes. Особенности строения. Бесполое размножение (многоспоровые и малоспоровые спорангии, спорангиоспоры, переход к конидиальному спороношению). Половое размножение (примитивная гаметангиогамия = зигогамия). Гаплофазный цикл развития. Представление о зигомицетах как о полифилетической группе.
Порядок Mucorales. Основные представители, особенности размножения. Экологические особенности и практическое значение представителей порядка.
Порядок Entomophthorales Особенности строения и размножения. Экологические особенности и практическое значение представителей порядка
Отдел Glomeromycota (порядок Glomerales) как одна из наиболее древних групп грибов. Везикулярно-арбускулярная микориза и ее роль в природе.
Царство Mycota
Отдел Zygomycota
Класс Zygomycetes
Порядок Mucorales : Mucor, Rhizopus, Syzygites (=Sporodinia)
Порядок Entomophthorales : Entomophthora
Отдел Glomeromycota
Порядок Glomerales : Glomus
Общая характеристика:
1. Вегетативное тело – неклеточный многоядерный мицелий, реже многоклеточный, септы с центральной долипорой. Есть мицелиально-дрожжевой диморфизм: В одних условиях они могут существовать в виде мицелия, в других – как отдельные почкующиеся клетки.
2. В кл. стенках хитин и хитозан.
3. При бесполом размножении образуются либо спорангии на спорангиеносцах со спорангиоспорами либо спорангии с небольшим числом спор - спорангиоли или конидии.
4. Половой процесс – зигогамия: концы двух гиф сближаются (гифы разного знака), на каждом конце отделяется по клетке, они сливаются между собой, зигота имеет характер покоящейся споры (зигоспора), прорастает (мейоз) в зародышевый спорангий (гаплонт).
5. Наземные сапротрофы, паразиты на грибах, насекомых и некоторых других животных.
Класс Zygomycetes
Подвижные стадии в цикле развития отсутствуют. Вегетативное тело – обильно разветвленный неклеточный многоядерный мицелий, субстратный и воздушный! В клеточной стенке – хитин в комплексе с хитозаном.
Бесполое размножение осуществляется неподвижными эндогенными спорангиоспорами, образующимися в спорангиях, или реже экзогенными конидиями. В ряде семейств можно проследить эволюцию бесполого размножения: видоизмененные специализированных структур, анаморф, от эндогеных спорангиоспор(Mucor) через малоспоровые спорангии(Thamnidium ) к экзогенными конидиям (Сunninghamella). (т.к. наземная среда нестабильна, а конидии созревают по очереди).
Половой процесс: зигогамия. Слияние недифференцированного на гаметы содержимого двух клеток, отделяющихся перегородками от несущих их гиф. Зигоспора на суспензорах. После периода покоя происходит редукционное деление диплоидного ядра, и из зигоспоры вырастает короткая нить мицелия с зародошевым спорангием на конце!
Главным образом сапротрофы.
Порядок Mucorales : Mucor, Rhizopus, Syzygites (=Sporodinia)
Mucor: спорангиеносцы простые или ветвящиеся, спорангии более мнеее шаровидные, зиготы голые. Rhizopus: образование воздушных столонов, на них, на месте соприкосновения с субстратом, образуются пучки ризоидов и группы спорангиеносцев, спорангии шаровидные. Весьма обычная плесень. У Thamnidiuma – односпоровые спорангии, называемые спорангиолями.
У Pilobolus – аппарат отбрасывания. Развивается на помете скота. Спорангиеносцы растут вверх от вздутой клетки – трофоцисты. Развивается тургорное давление, кольцевой взрыв, спорангий выбрасывается до 1,8 метров в высоту и 3, 5 в длину.
Рассмотрим гаплоидный цикл у представителей отдела Zygomycota: Mucor sp. Мицелии многоядерные, но обязательно гаплоидные. С ухудшением условий среды гриб переходит в стадию полового размножения. Ищет себе партнера, обладающего противоположным знаком. Происходит отделение перегородкой от основного мицелия участка(многоядерного, гаплоидного), который и будет принимать участие в половом процессе. При романтическом соприкосновении данных участков от разных мицелиев происходит плазмогамия. В результате образуется многоядерная зооспора, обладающая разнокачественными ядрами(гетерокариоз). Зооспора держится на суспензерах. Внутри происходит происходит кариогамия. Образуется диплоидная зигоспора(хотя в результате слияния ядер часть может погибнуть). Мейоз при прорастании зиготы. Образуется зародышевый спорангий. Гриб вступает в стадию бесполого размножения(спорангиоспоры).
Порядок Entomophthorales : Entomophthora
Неклеточный мицелий – поражение насекомых. Появление перегородок и слияние именно «отдельных» участков. При бесполом – конидии, активно отстреливаются! Большинство видов порядка являются патогенными грибами насекомых. Некоторые виды поражают нематод, клещей и тихоходок, а также некоторые (в частности виды рода Conidiobolus) — сапротрофы. половой процесс – зигогамия.
Entomophthora muscae – вызывает гибель комнатных мух «осенняя болезнь мух». Мицелий многоклеточный, а сами клетки многоядерные. Когда конидия попадает в муху, прорастает в ростковую трубку, гифу, внедряющуюся в тело насекомого. В жировом теле мухи гифа разрастается и затем распадается на многоядерные клетки – «гифенные тела», которые разносятся по всему телу насекомого. Из дыхательных отверстий её хитинового покрова высовываются неветвящиеся булавовидные конидиеносцы, несущие на конце шаровидную многоядерную конидию. Проявляют довольно узкую специализацию к насекомым-хозяинам.
Отдел Glomeromycota:
Порядок Glomerales : Glomus
Образуют наиболее распространенный тип микориз – везикулярно-арбускулярные микоризы (ВАМ). Причем даже с такими растениями, как злаки. Одна из наиболее древних групп грибов. Все представители – возбудители микоризообразования. Подовляющее большинство с травенистыми растениями. Почвенные грибы.
Морфология: эндомикориза – внутри корня располагается большая часть. Арбускулы – части гиф, входящие в клетку хозяина. Формирование внутри везикул – увеличенные клетки с утолщенной стенкой с большим запасом питательных веществ.
При размножении – образуются споры, вроде бы, так как у специалистов ещё нету определенной точки зрения. Образуются сплетения спор и гиф (спорокарпы), сами споры – очень сложно устроенная клеточная стенка(переплитания и несколько слоев).