

61 Общая схема энергетического обмена гетеротрофов.
Все живые организмы и микроорганизмы используют в кач-ве ист. Е 3 основных источника:1) это Е ЭМ излучения, 2) для литотрофов(неорганотрофов и хемосинтетиков) – реакции окисления неорг. соединений 3)для гетеротрофов – р-ции окисления органических субстратов. Е обмен бактерий и др.живых организмов, у кот.он присутствует,выполняет такие ф-ции: 1)трансформация Е внешних энергетич. ресурсов в источник конвертированной Е,2)обеспечение этой Е всех видов работы в клетке.
hν- Е кванта ЭМ излучения – характерно для: высших р-ний, цианобактерий, водорослей, некоторых бактерий. Схема транслокации Е у прокариотов. Пояснение схемы. О.Н.С(окисление неорг.соед.)-хар. Для литотрофов и органотрофов. О.О.С(ок.орг.соед.) – гетеротрофы по ист.Е: высшие животные, человек, грибы, подавл. большинство простейших.
Конвертируемая Е – это Е, которая может быть преобразована, использована для работы.
Сопрягающие мембраны – те мембраны, которые формируют стр-ры, отвечающие за трансформацию Е клетки. У животных это внутренняя митохондр. мембрана. У растений – внутр. мембр. хлоропластов, у цианобакт. – фикобилисома, у бактерий – процессы локализованы в цитопл. мембране.
Х.Р. – химическая работа – это весь конструктивный обмен, О.Р. – осмотическая работа(1) поддерж. Клеточного гомеостаза, 2)активный транспорт в-в) Э.Е. – электрич. энергия дополнительная(это значит, что часть Е у бактерий может превращаться в световую – хемобиолюминесценция – способны выд. морские и др. водеые бактерии)Т.Э. – тепловая энтропия. Нужно помнить, что живые организмы умеют использовать Е только электронов, но не атомов.
Е внешних энергет. источников переходит в ист. конвертируемой Е. У бактерий – з типа источников конв.Е: 1)электрохимич. потенциал ионов водорода ΔμН, который образуется в процессе работы двхательной цепи или редокс-цепи фотосинтетического аппарата. 2) АТФ, кот. образуется во время субстратного, окислительного или фотосинтетического фосфорелирования.3) дополнительный фонд макроэргов – неорг. полифосфаты это полимеры, кот. состоят из остатков неорг. фосфата, соединенных между собой макроэргическими связями. Все эти источники могут трансформироваться 1 в другой, в зависимости от условий сущ-вования и функционального состояния клетки. Транвформация Е АТФ и наоборот зависит от типа работы, которую выполняет клетка. При интенсивном уровне биосинтеза необходима Е АТФ и некоторых других соединений: гуанинтрифосфата(ГТФ), урацилтрифосфата(УТФ), цитозинтрифосфат(ЦТФ). В случае интенсификации систем активного транспорта наиболее требуемой становится Е ΔμН.
Вместо всего этого – схема из конспекта (общая схема ен обмена гетерофов)
62 Характеристика метаболического пути по схеме Энтнера-Дудорова.
Этот путь получил название 2-кето-3-дезокси-6-фосфоглюконатного (КДФГ) пути расщепления углерода.
Первые два этапа- фосфорилированние молекулы глюкозы и ее дегидрирование до 6-фосфоглюконовой кислоты – идентичен первым двум этапам ГМФ-пути. Специфичны для Энтера-Дудорова две следующие реакции: дегидратирование 6-фосфоглюконовой кислоты, катализируемое 6-фосффоглюканат-дегидратазой и приводящая к образованию КДФГ – кислоты; расщепления продукта первой реакции а два С3-фрагмента с помощью фрагмента КДФГ-альдозы.
Конечным продуктом второй реакции являются пировиноградная кислота и 3-ФГА. Последний окисляется в пировиноградную кислоту так же, как в гликолизе. Следовательно при разложении одно молекулы глюкозы образуется одна молекула АТФ.
Этот путь характерен для рода Pseudomonas.
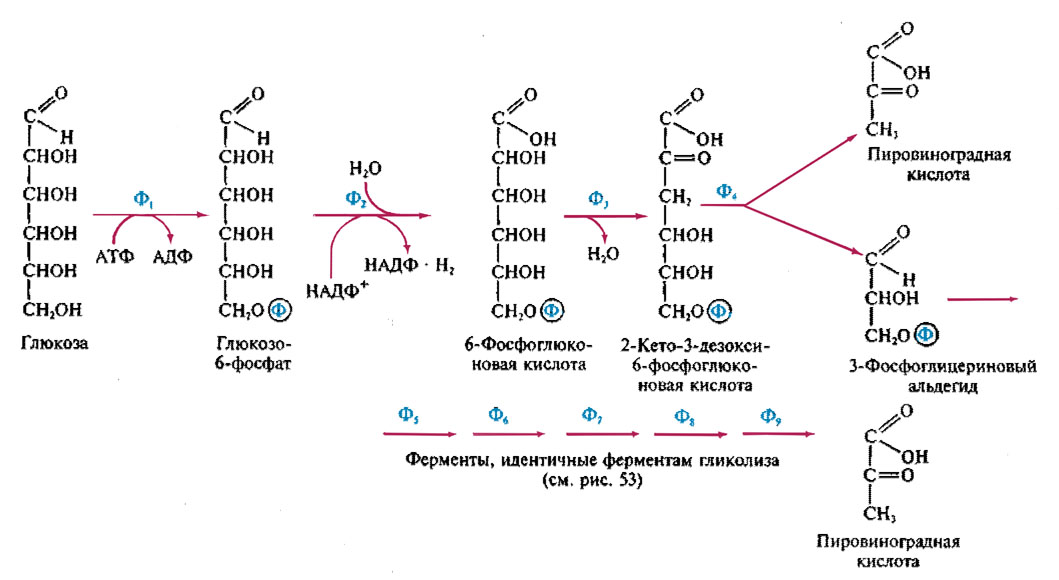 Ф1
- гексокиназа; Ф2 - глюкозо-6-фосфатдегидрогеназа;
Ф3 - 6-фосфоглюконат-дегидратаза; Ф4 -
2-кето-3-дезокси-6-фосфонат- алолаза; Ф5 -
глицеральдегид-3-фосфатдегидрогеназа;
Ф6 - фосфоглицераткиназа; Ф7 -
фосфоглицеромутаза; Ф8 - фосфоглицеромутаза;
Ф9 - пируваткиназа.
Ф1
- гексокиназа; Ф2 - глюкозо-6-фосфатдегидрогеназа;
Ф3 - 6-фосфоглюконат-дегидратаза; Ф4 -
2-кето-3-дезокси-6-фосфонат- алолаза; Ф5 -
глицеральдегид-3-фосфатдегидрогеназа;
Ф6 - фосфоглицераткиназа; Ф7 -
фосфоглицеромутаза; Ф8 - фосфоглицеромутаза;
Ф9 - пируваткиназа.
63 . Генетичні та біохімічні основи антибіотико резистентності. Шляхи подолання.
Многие антибиотики утратили свою эффективность в святи с развитием устойчивости мо к ним. Может бать 2-х типов: Естественная – обусловлена отсутствием мишени для АБ. Такого звена, которое блокировалось под влиянием препарата. Приобретенная - обусловлена мутаціями в генах, контролирующих синтез ЦПМ, белков либо возникает в результате генетических рекомбенаций в ходе переноса плазми (R – фактора), контролирующих множественную устойчивость к АБ. Плазмиды включают комплекс генов, кодирующих синтез ферментов, разрушающих либо модифицирующих структуру АБ. Биохимические основы антибиотикорезистентности. Устойчивость мо связана с изменением обменных процессов в микробной клетке: 1) Устойчивые мо синтезируют ферменты, приводящие антибиотик в неактивную форму путем модификации или инактивации; 2) Модификация мишени на которую действуют АБ путем инактивации рецепторов; 3) Изменение проницаемости Кл → непроницаема для АБ (изменение свойс белка переносчика); 4) Более интенсивное образование фермента на который действует АБ. 5) активне виведення антибіотика з клітини (ефлюкс).Для преодоления лекарственной резестентности бактерий необходимо пополнение арсеналахимтерапевтических препаратов новыми, которые отличались новыми механизмами антимикробного действия. Перспективным является химическая модификация уже известных АБ, изыскание ингибиторов ферментов инактивирующий АБ.
Генетично резистентність може кодуватися хромосомними генами (хромосомна стійкість) або генами, шо мають плазмідну локалізацію (позахромосомиа, плазмідна резистентність). Плазміди, шо кодують резистентність, називають R-плазмідами.
Генетичні детермінанти антибіотикорезистентності можуть передаватися від однієї клітини до іншої в ході кон'югації, трансдукції й трансформації. Антибіотикостійкість може бути результатом мутацій, що приводять до модифікації мішені дії антибіотика або інших структурно-функціональних змін у клітині.