46 Cинтез нуклеотидов у бактерий.
Нуклеиновые кислоты представляют линейные полимеры нуклеозидмонофосфатов, то есть полинуклеотиды. Нуклеотиды построены из трех компонентов: пиримидинового или пуринового основания, пентозы и фосфорной кислоты.
Синтез пуринов и пиримидинов отличается. В подавляющем большинстве м/о синтезирут нуклеотиды самостоятельно. В основе пуринов лежит рибоза-5-Р, соединяясь с АТФ – 5-Р-рибозил-1-пирофосфат (фрпф). К нему присоединяются сахара, ГЛИ, NH3 CO2 Образуется инозин монофосфат.
(СХЕМА ВЕРШИГОРА стр 167)
47 Цикл Арнона у автотрофов
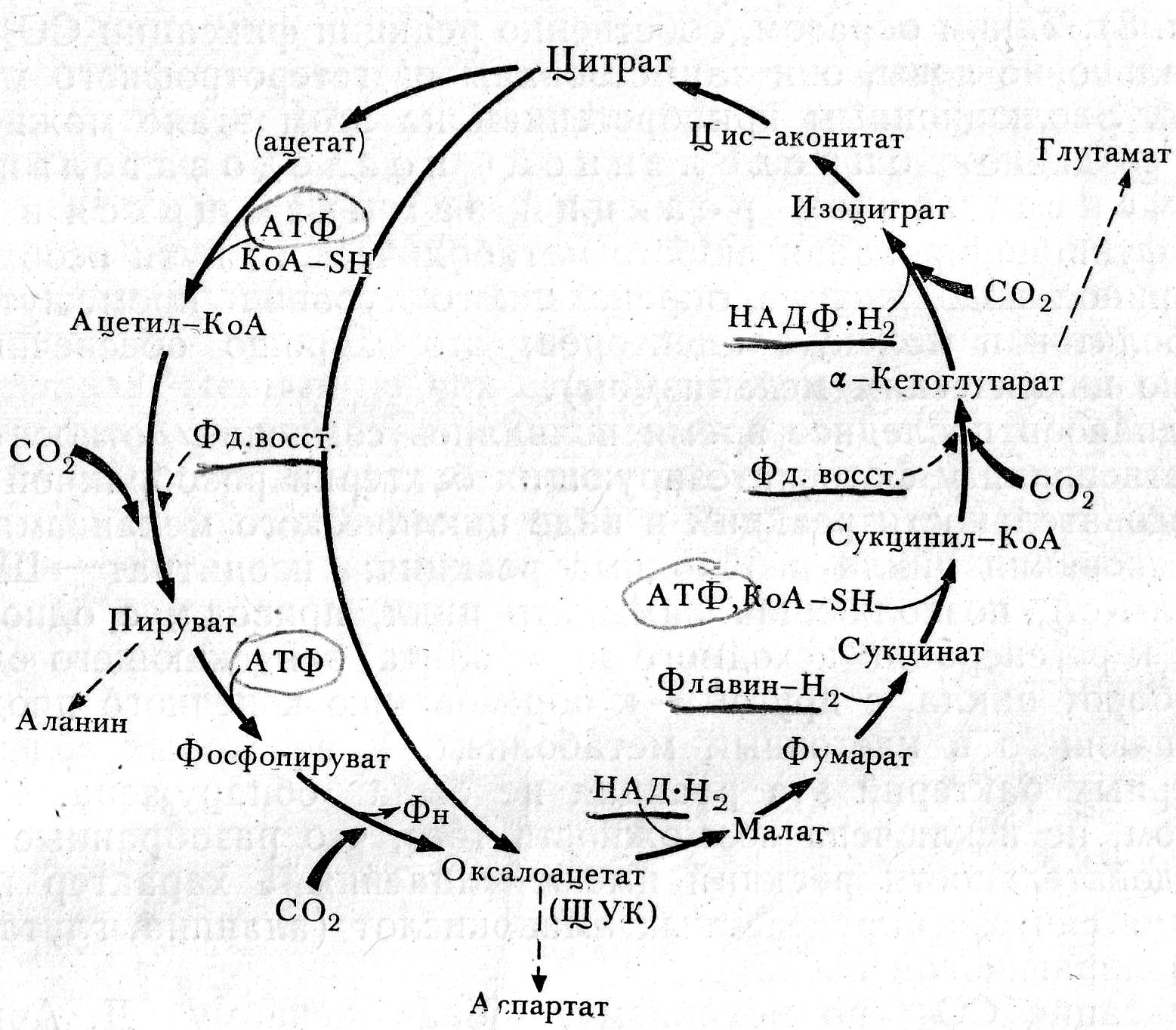
Цикл Арнона эволюционно более древний путь фиксации СО2, чем цикл Кальвина, имеет реакции ЦТК в обратном направлении. Основа фиксации СО2 – восстановительное карбоксилирование органических кислот. СО2 в цикле фиксируется в 4-х ферментат реакциях, две из которых идут при участии фотохимически восстановленного ферредоксина. В результате одного цикла из 4-х молекул СО2, 10[H] и энергии 3-х молекул АТФ синтезируется конечный продукт – оксалоацетат (щавелевоуксусная кислота, или ЩУК) – конечный продукт цикла.
Цикл Арнона не имеет широкого распространения у фотосинтезирующих прокариот. Обнаружен у зелёных серобактерий из рода Chlorobium и у некот пурпурных бактерий

48 Синтез жк и глицерина
Жирные кислоты метаболизируются через каскад окислительных реакций. Цель этих превращений — образование ацетил-КоА, вступающего в цикл Крёбса. Первичные продукты фотосинтеза, хемосинтеза и образовавшиеся из них или поглощённые углеводы являются исходным материалом для синтеза липидов — жиров и др. жироподобных веществ. Некоторые микроорганизмы (например, Torulopsis lipofera) при культивировании на растворах глюкозы за 5 часов образуют до 11% жира на сухое вещество. Глицерин, необходимый для синтеза жиров, образуется путём восстановления фосфоглицеринового альдегида. Высокомолекулярные жирные кислоты — пальмитиновая, стеариновая, олеиновая и др., дающие при взаимодействии с глицерином жиры, синтезируются в организме из уксусной кислоты — продукта фотосинтеза или окисления веществ, образовавшихся в результате распада углеводов.
49
50 Механизм транспорта питательных веществ в бактериальные клетки.
Механизм транспорта питательных веществ в бактериальную клетку.Существует 4 типа транспорта питательных веществ в Бак. Кл.: 1) Простая (пассивная) диффузия неспецифические проникновения вещества в клетки по градиенту химического или электрохимического потенциала без затраты энергии до уравнивания концентрации по обе стороны ЦПМ. По такому механизму в бакт.кл. транспорт Н2О, ионы, ингибитори2) Облегченная диффузия-транспорт веществ в комплексе с белковым переносчиком-пермеаз. Энергия расходуется только на синтез переносчика. Проникновение веществ в кл. происходит по градиенту химического или электрохимического потенциала в уравнении концентрации по обеим сторонам ЦПМ 3) Активный транспорт - это транспорт веществ против градиента хим. потенциала, на который расходуется большое количество энергии. Процесс катализируется перемеазами.впитываться могут только вещества для которых существуют транспортные системи.4) Активный транспорт с модификациями субстрата-транслоация радикалив.Транспорт только одного вещества по градиенту электрохимического трансмембранным потенциала назыв. унипортом. Если белок катализирует одновременно и направленых перенос одного протона и одной молекулы субстратов, то речь идет о симпорт 2-х или нескольких веществ, другие белки канализируют одновременное переноса в противоположных направлениях двух 2 частиц, например протонов-ионов Na.У таких случаях говорят о антипорте. В прокариот преобладает симпорт веществ с протонами
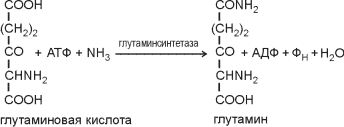
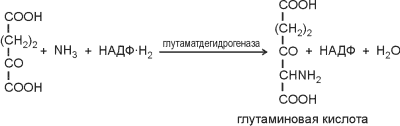
51 Синтез аминокислот у бактерий. Большинство бактерий самостоятельно синтезируют все 20 аминокислот, в отличие от животных. По своим предшественникам аминокислоты делятся на 5 групп, в них не входит только гис.
-
глутаминовая
Цтк
альфакетоглутарат и ион аммония
глу кта,глу. (арг,про-2 перестройки С цепей, переаминирование)
аспарагиновая
Цтк
ЩУК
асп к-та (переаминирования ЩУК), асп (переаминирование асп кты), лиз,мет,тре,изолей
ароматическая
гликолиз, варбурга-диккенса-хорегера
фосфоенолпвк+эритозо4фосфат
три,тер,фен
сериновая
интермедиат энтнера дудорова
3-Р-глицерат
сер, из него цис, гли
группа ПВК
гликолиз, энтнера дудорова
ПВК
ала (пвк+глутамат - ала+альфакетаглутарат),вал,лей
гистидин
Кальвина
5-Фосфорибозил-1-пирофосфат
5-Фосфорибозил-1-пирофосфат+АТФ+N+ГЛУ=гис