

Химические свойства.
Оксокислоты вступают в реакции, характерные для карбоксильной и карбонильной групп. Отличительная черта оксокислот – легкость, с которой протекает их декарбоксилирование.
a -Оксокислоты легко отщепляют СO2 и СО при нагревании в присутствии серной кислоты.
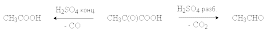
b –Оксокислоты неустойчивы и самопроизвольно декарбоксилируются с образованием кетонов.
CH3COCH2COOH ® CH3COCH3 + CO2
b -Оксокислоты и их эфиры обладают специфическими свойствами, которые связаны с их повышенной
СН-кислотностью. Повышенная подвижность протонов метиленовой группы обусловлена
электроноакцепторным влиянием двух карбонильных групп. В результате b -оксокислоты существуют в виде двух таутомерных форм: кетонной и енольной (см. лек. №11), причем содержание енольной формы в равновесной смеси значительное. Енольные формы дополнительно стабилизируются за счет наличия в них системы сопряженных p -связей и внутримолекулярной водородной связи.

При действии на ацетоуксусный эфир какого-либо реагента в реакцию вступает один из таутомеров. Поскольку второй таутомер за счет смешения равновесия восполняет убыль первого, таутомерная смесь реагирует в данном направлении как единое целое.
19.Химический синтез аминокислот. Способы разделения рецематов. Понятие о ферментативном и микробиологическрм синтезе.
Синтез аминокислот.Если в недавнем прошлом -аминокислоты получали в небольших количествах, преимущественно для научных исследований, то в настоящее время налажено их многотоннажное промышленное производство. Это связано, например, с тем, что -аминокислоты являются необходимым компонентом комбикормов и синтетической пищи на углеводной основе.
Химический синтез аминокислот.
В лабораторных условиях химический синтез аминокислот осуществляют при взаимодействии
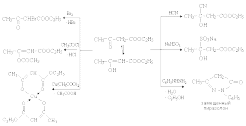
-галогенокарбоновых кислот с аммиаком. Исходные -галогенокислоты обычно получают по реакции Гелля–Фольгарда–Зелинского:
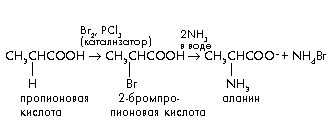
В синтезе Штреккера альдегид превращают в -аминокислоту с удлинением углеродной цепи на один атом углерода. Процесс проходит в две стадии. На первой стадии в результате реакции альдегида с NH4Cl и NaCN получают -аминонитрил; на второй – при гидролизе нитрильной группы -аминонитрила получают аминокислоту:

Очень распространен способ получения -аминокислот с использованием малонового эфира. При этом один из атомов водорода метиленовой группы малонового эфира замещается на аминогруппу, а другой – на соответствующий требуемой аминокислоте углеводородный радикал. В результате гидролиза полученного диэтилового эфира и последующего декарбоксилирования дикислоты получают нужную аминокислоту:

Еще один способ синтеза аминокислот заключается в восстановительном аминировании (восстановлении водородом в присутствии аммиака) -оксокарбоновых кислот:

Наиб. общий метод расщепления рацематов-химический, при к-ром на рацематы действуют оптически активным реагентом, в результате чего образуется новая пара в-в - диастереоме-ров. Последние м. б. разделены вследствие различия в их физ. св-вах. Хиральный реагент после разделения диасте-реомеров отщепляют. Напр., рацемич. (R, S)-1-фенилэтил-амин образует с природной (2R, 3R)-винной к-той две диасте-реомерные соли: [(R)-1-фенилэтиламин]·[(2R,3R)-винная к-та] и [(S)-1-фенилэтиламин]·[(2R, 3R)-винная к-та], к-рые обладают разл. р-римостью в этаноле и м. б. разделены кристаллизацией. Своб. амин выделяют затем экстракцией диэтиловым эфиром из водного щелочного р-ра соли.
Для расщепления рацемич. к-т используют их способность образовывать соли с хиральными прир. и синтетич. основаниями-хинином, бруцином, стрихнином, 1-фенилэтилами-ном и др. Рацемич. спирты расщепляют путем превращения их в кислые эфиры дикарбоновых к-т (напр., фталевой) с послед. кристаллизацией в виде диастереомерной соли с хиральным амином.
Метод расщепления рацематов путем превращения их в диастереомеры не пригоден для орг. соед., не имеющих функц. групп, напр. для алканов. Для расщепления таких рацематов используют, напр., способность мочевины к образованию клатратов. Мочевина кристаллизуется в хиральной гексагон. решетке, в цилиндрич. каналах к-рой могут размещаться молекулы "гостя". Кристаллы мочевины м. б. как право-, так и левовращающими, поэтому клатраты мочевины с энантиомера-ми приобретают характер диастереомеров. Разделение рацематов по этому способу осуществляют внесением затравки одного из энантиомеров в насыщ. р-р мочевины и рацематов, выпадающие при этом кристаллы клатрата обогащены именно внесенным энантиомером.
Др. хим. метод расщепления рацематов-кинетич. расщепление, основанное на том, что в р-циях с оптически активными реагентами (или в присут. хиральных катализаторов или хиральных р-рителей) скорость превращения одного энантиомера не равна скорости превращения другого. Если в подобную р-цию ввести рацемат и прервать р-цию до ее полного завершения, то один из энантиомеров, реагируя быстрее, будет преобладать в продукте р-ции, другой-в непрореагировавшем остатке. Пример - расщепление рацемич. (n-толил) мезитилсульфоксида восстановлением под действием реагента, полученного из оптически активного 1-фенилэтиламина и А1Н3.
Асим. превращениями наз. процессы, в ходе к-рых происходит превращение рацематов в смесь энантиомеров с преобладанием одного из них. К асим. превращениям относится, в частности, мутаротация моносахаридов. Этот метод наз. также ретрорацемизацией.
Для расщепления рацематов используют также хроматографию на хиральных неподвижных фазах. Ранее в качестве таких фаз использовали прир. хиральные полимеры - белки, крахмал, целлюлозу, шерсть и др., из к-рых теперь применяют только микрокристаллич. целлюлозу и ее производные. В осн. для расщепления рацематов используют более селективные синтетич. хиральные сорбенты, полученные специально для разделения той или иной группы рацемич. соединений. Так, рацематы, молекулы к-рых содержат фрагменты, способные к образованию комплексов с переносом заряда, м. б. расщеплены на хиральных фазах, имеющих структуру нафтилглицина (для электроноакцепторных молекул) или динитробензоил-аминокислот и пикрил (1-фенилэтил) амина (для электроно-донорных молекул). Соед., способные к образованию комплексов с переходными металлами, м. б. разделены на энантиомеры методом лигандообменной хроматографии с использованием хиральных комплексообразующих сорбентов. Этим методом расщепляют, напр., меченные тритием a-аминокислоты. Применяют сорбенты, содержащие фрагменты циклодекстринов, расщепление рацематов на них осуществляется в результате образования соед. включения. Получены хиральные стационарные фазы, разделение энантиомеров на к-рых происходит благодаря возникновению водородных связей и ион-дипольных взаимод. между энантиомерами и сорбентом. Расщепление рацематов хроматографич. методом позволяет одновременно получить и информацию об оптич. чистоте выделенных энантиомеров.
Микробиологический синтез- синтез структурных элементов или продуктов обмена веществ микроорганизмов за счёт присущих микробной клетке ферментных систем. При М. с., как и любом органическом синтезе, сложные вещества образуются из более простых соединений. М. с. следует отличать от брожения (См. Брожение), в результате которого тоже получаются различные продукты микробного обмена (например, спирты, органические кислоты), но преимущественно за счёт распада органического вещества. Значительная часть продуктов, образующихся в ходе М. с., обладает физиологической активностью и представляет практическую ценность для народного хозяйства.
К М. с. относят широкий круг процессов. 1. Накопление микробной массы для использования её: а) в качестве белково-витаминных добавок к кормам; б) как источника получения белков (См. Белки), липидов (См. Липиды), ферментов (См. Ферменты), токсинов (См. Токсины), витаминов (См. Витамины), антибиотиков (См. Антибиотики); в) для борьбы с паразитами животных и растений; г) в качестве носителя ферментативной активности в реакциях микробиологической (энзиматической) трансформации органических соединений. 2. Получение накапливающихся вне микробной клетки метаболитов, в том числе ферментов, токсинов, антибиотиков, аминокислот, витаминов, нуклеотидов и т.п.
М. с. осуществляется внутри клетки при активации низкомолекулярных компонентов (например, коферментом А (См. Кофермент А)) и участии нуклеотид фосфатов, чаще всего адениловых производных (см. Аденозинфосфорные кислоты). Затем многие метаболиты выводятся из клетки в среду. Характерная особенность микроорганизмов — их способность к сверхсинтезу, т. е. избыточному образованию некоторых продуктов обмена веществ (многих аминокислот, нуклеотидов, витаминов), превышающему потребность микробной клетки. Так, глутаминовая кислота при сверхсинтезе может накапливаться в количестве свыше 10 мг/мл среды (культура Micrococcus glutamicus), витамин B2 — до 1—2 мг/мл (грибы Eremothecium ashbyii u Ashbya gossipii), вместо обычных сотых и даже тысячных долей мг. Способность к сверхсинтезу того или иного соединения свойственна определённым видам микроорганизмов, которыми, как правило, и пользуются в качестве продуцентов при производстве соответстветствующих метаболитов путём М. с. При этом применяют не только культуры, отобранные из природных источников, но и специально выведенные искусственным путём Мутанты — штаммы, у которых сверхсинтез — следствие нарушений обмена веществ под воздействием мутагенов (См. Мутагены). Применение мутантов позволяет значительно увеличить выход ряда продуктов. Например, выведены культуры с высоким уровнем сверхсинтеза лизина, инозиновой кислоты, некоторых витаминов. При помощи мутантов удалось в 100—150 раз поднять активность биосинтеза пенициллина (См. Пенициллины); мутантные штаммы используются при производстве как этого, так и др. антибиотиков.
В процессе М. с. получают ряд продуктов, причём за счёт самых разных соединений углерода и азота. Это обусловливается большим разнообразием ферментных систем микроорганизмов. Так, для синтеза белков, нуклеиновых кислот и др. метаболитов клетки могут использовать в зависимости от особенностей культуры разные неорганические источники азота, а из соединений углерода — различные углеводы, органические кислоты (в т. ч. уксусную кислоту), жидкие, твёрдые или газообразные углеводороды и др. Определённые виды, способные к Хемосинтезу или Фотосинтезу, в качестве источника углерода могут усваивать углекислый газ. Т. о., подбор соответствующих культур даёт возможность получать путём М. с. желаемые вещества из дешёвого и доступного сырья. Эти особенности делают М. с. весьма эффективным способом производства многих соединений; часть из них (например, многие антибиотики) экономически выгодно получать ныне только таким путём.
Некоторые продукты М. с. давно использовались человеком (например, пекарские дрожжи), но широкое промышленное применение М. с. получил начиная с 40—50-х гг. 20 в. Прогресс в этой области связан прежде всего с открытием пенициллина, что побудило начать детальные исследования у микроорганизмов продуктов обмена веществ, обладающих физиологической активностью. Освоение в промышленных масштабах производства пенициллина привело к решению многих микробиологических, технологических и инженерных задач. Это, наряду с расширением производства дрожжей как белково-витаминных добавок к кормам, послужило основой для развития промышленного М. с. Так, в частности, были созданы специальные аппараты — ферментёры, с помощью которых можно вести технологический процесс биосинтеза без доступа посторонних микроорганизмов, снабжённые устройствами для перемешивания среды и для подачи стерильного воздуха.
Под ферментативным синтезом гена имеют в виду транскрибирование комплементарной нити ДНК (гена) на молекулах РНК в пробирке. Система для синтеза представляет собой раствор, в котором содержатся все четыре нуклеотида, входящих в состав ДНК, ионы магния, фермент обратная транскриптаза (ее получают из онкогенных вирусов) и матричная (информационная) РНК, кодированная геном, копию которого ставится задача снять. На иРНК обратная транскриптаза синтезирует комплементарную ей цепь ДНК, а затем на ней при помощи этого же фермента синтезируется вторая цепь ДНК. В результате получается ген по структуре такой же, как и тот, на котором была синтезирована иРНК.
Этим способом в лабораториях многих стран создан целыйряд генов. В нашей стране йод руководством В. А. Энгельгардта был разработан проект «Ревертаза» — программа синтеза генов с помощью этого фермента. В осуществлении проекта участвовали ведущие отечественные и зарубежные институты. В итоге с 1974 по 1978 г. были синтезированы гены глобина голубя, кролика и человека, а также гены митохондрий печени крыс, часть гена, кодирующего иммунные белки мышей, и др.