

Гипотетические механизмы тахифилаксии при эпидуральном введении локальных анестетиков
Гипотезы |
Публикации |
Фармакокинетические |
|
Ограничение диффузии из-за периневрального отека |
61 |
Подкисление ЦСЖ, эпидурального пространства, невральных тканей, снижающее количество липофильных форм анестетика |
57, 59 |
Усиленная элиминация или нарушения распределения локального анестетика |
60, 61 |
Увеличение концентрации натрия в эпидуральном пространстве, вследствие чего диффузия анестетика снижается |
54 |
Фармакодинамические |
|
Увеличение содержания циклического аминомонофосфата |
62 |
Нейропластические |
|
Усиление ноцицептивной трансмиссии, периферическая и центральная сенсибилизация ноцицепторов |
47, 52, 68 |
Величина рН. Повторные инъекции коммерческих растворов локальных анестетиков вызывают ацидификацию ЦСЖ и невральных тканей [57]. Повышение концентрации ионов водорода в эпидуральном пространстве и ЦСЖ (снижение рН) сопровождается увеличением доли заряженных анионных форм локального анестетика по сравнению с основной неионизированной формой. Поскольку липофильной является только неионизированная форма локального анестетика, то с уменьшением ее доли по мере падения рН снижается и диффузия препарата через нервную мембрану. Слабая буферная способность ЦСЖ (и, возможно, эпидурального пространства) делает эти среды особенно восприимчивыми к ацидификации [58].
Исследования последних лет не подтвердили гипотезу ацидификации. Растворы бупивакаина со стабильным рН 4,2–6,8 повторно вводили с помощью имплантированной системы в область седалищного нерва крысы, омывая его. Несмотря на 400-кратное различие в концентрации водородных ионов, никакой разницы в развитии тахифилаксии не отмечено [59].
Усиленное выделение или нарушение распределения локальных анестетиков. В последнее время важное значение в развитии тахифилаксии стали придавать усиленной элиминации бупивакаина из эпидурального пространства. Mogensten и сотр. [60], изучая клиренс 133Хе, установили, что после развития эпидуральной блокады 0,5% раствором бупивакаина кровоток в эпидуральном пространстве усиливался. Усиление этого кровотока продолжалось и в последующем у пациентов, уровень сенсорной анестезии у которых снижался в процессе продолжительной инфузии бупивакаина, тогда как у пациентов со стабильным уровнем сенсорной анестезии дальнейшего нарастания эпидурального кровотока не происходило [60]. В то же время усиленная элиминация анестетика из эпидурального пространства или изменения его распределения в этом пространстве не играли роли при развитии тахифилаксии на фоне интер-миттирующих инъекций лидокаина [61].
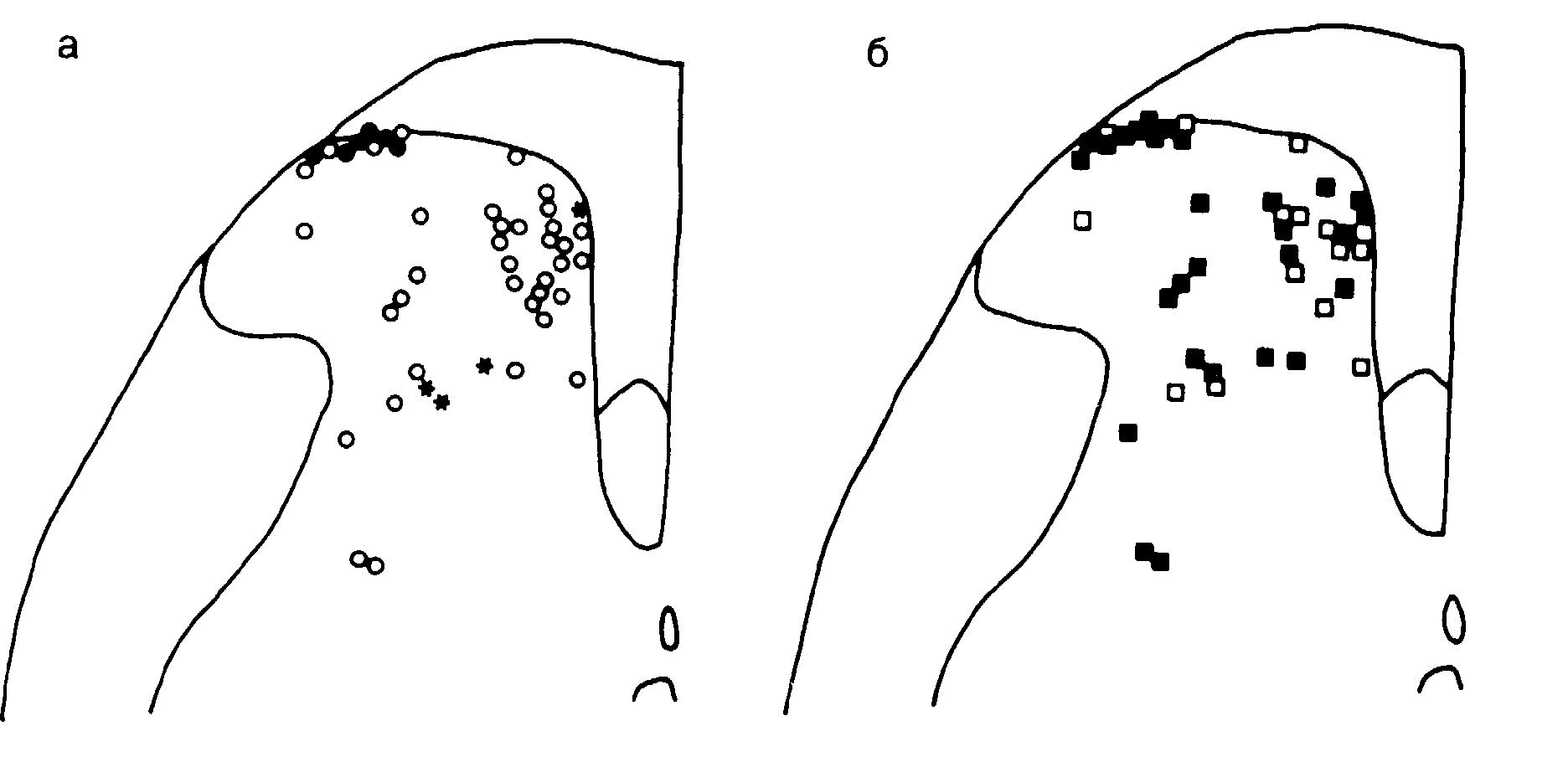
Рис. 12-4. Разрез заднего столба спинного мозга. Распространение рецептивных полей нейронов заднего рога.
На поперечном сечении заднего рога спинного мозга крысы проверено расположение 46 нейронов, а-клетки, активированные после стимуляции дорсолатерального пучка на противоположной стороне спинного мозга (черные кружки) и вентролатерального квадранта (звездочки); некоторые клетки остались нсстимулиро ванными (светлые кружочки); стимуляцию проводили вольфрамовым электродом; б клетки, расширившие свои рецептивные поля более чем на 50% или усилившие свою реакцию на стандартный щипок либо на стимул с форсункой, после обработки С-волокон (черный квадрат); клетки, оставшиеся несенсибилизированными при обработке (светлый квадрат). (По Cook и др. [65], с разрешения.)
Фармако динамические гипотезы. Увеличение цАМР. Недавние исследования in vivo и in vitro, касающиеся блокады седалищного нерва прокаином, показали, что действие локальных анестетиков (включая тахифилаксию), возможно, опосредовано аденином и циклическими нуклеоти-дами [62]. Вещества этих групп значительно сокращают продолжительность невральной блокады у крыс, не влияя при этом на частоту, степень выраженности блокады и время ее наступления. Под влиянием нуклеотидов происходит быстрое восстановление потенциала действия изолированного нерва, подавленного прокаином. Необходимы дальнейшие исследования роли аденина и циклических нуклеотидов в генезе тахифилаксии и в других аспектах действия локальных анестетиков.
Усиленная ноцицептивная трансмиссия и нейропластичность. Наиболее интригующая гипотеза о механизме тахифилаксии основана на физиологии ноцицептивной сенсорной системы [52]. Лабораторные эксперименты показали, что после периферической травмы усиливаются ноцицептивная трансмиссия, периферическая и центральная сенсибилизация ноцицепторов [63–66] (гл. 2). При подобном увеличении афферентных импульсов, поступающих в спинной мозг, рецептивные поля нейронов заднего рога расширяются [65] (рис. 12-4 и 12-5). Наряду с этим происходит прогрессирующая гиперсенсибилизация нейронов заднего рога (рис. 12-6). Примечательно, что повреждающая стимуляция глубоких соматических структур выявляет более значительное ноцицептивное заграждение и более выраженные центральные нейропластические изменения, чем повреждающая стимуляция кожных структур [67]. Экстраполируя эти данные, можно полагать, что повреждающая стимуляция более глубоких висцеральных структур также вызывает более значительное афферентное заграждение, чем кожная стимуляция.
Подобные же изменения ноцицептивной трансмиссии и центральной гиперсенсибилизации могут иметь место и в послеоперационном периоде [68]. Усиленный поток афферентных импульсов и перевозбуждение нейронов заднего рога вступают в антагонизм с блокирующим действием эпидурально введенных анестетиков. Данный антагонизм будет проявляться сам по себе как редукция сенсорной аналгезии на уровне рострального отдела невральной блокады, где интенсивность блокады и содержание бупивакаина наименьшие [47, 52, 68]. Особую неотразимость данной гипотезе придает следующий вывод: ноцицептивная трансмиссия и сопровождающие ее нейропластические изменения могут быть устранены комбинацией анальгетиков, действующих на дискретные отделы ноцицептивной системы (т. е. с помощью балансированной аналгезии).

Рис. 12-5. Распространение рецептивных полей заднего рога.
Изменение размеров рецептивных полей нейронов заднего рога под воздействием обрабатывающего стимула определенной силы, направленного на икроножный нерв крысы. Заштрихованные участки обозначают кожные зоны, при нежном механическом раздражении которых выявляется реакция нейрона, а – действие стимулов, активирующих только большие миели-низированные афферентные волокна (100 мкА, 50 мкс); б – влияние наиболее сильного стимула на вовлечение тонких миелинизированных афферентных волокон (500 мкА, 50 мкс): в – эффект, возникающий при стимулировании немиелинизированных волокон (5 мА, 500 мкс). Отмечено расширение рецептивных полей нейронов заднего рога. Цифры рядом с каждой лапкой- время (мин), обрабатывающий стимул накладывался во время 0 при 1 Гц на 20 с. (По Cook и сотр. [65], с разрешения.)
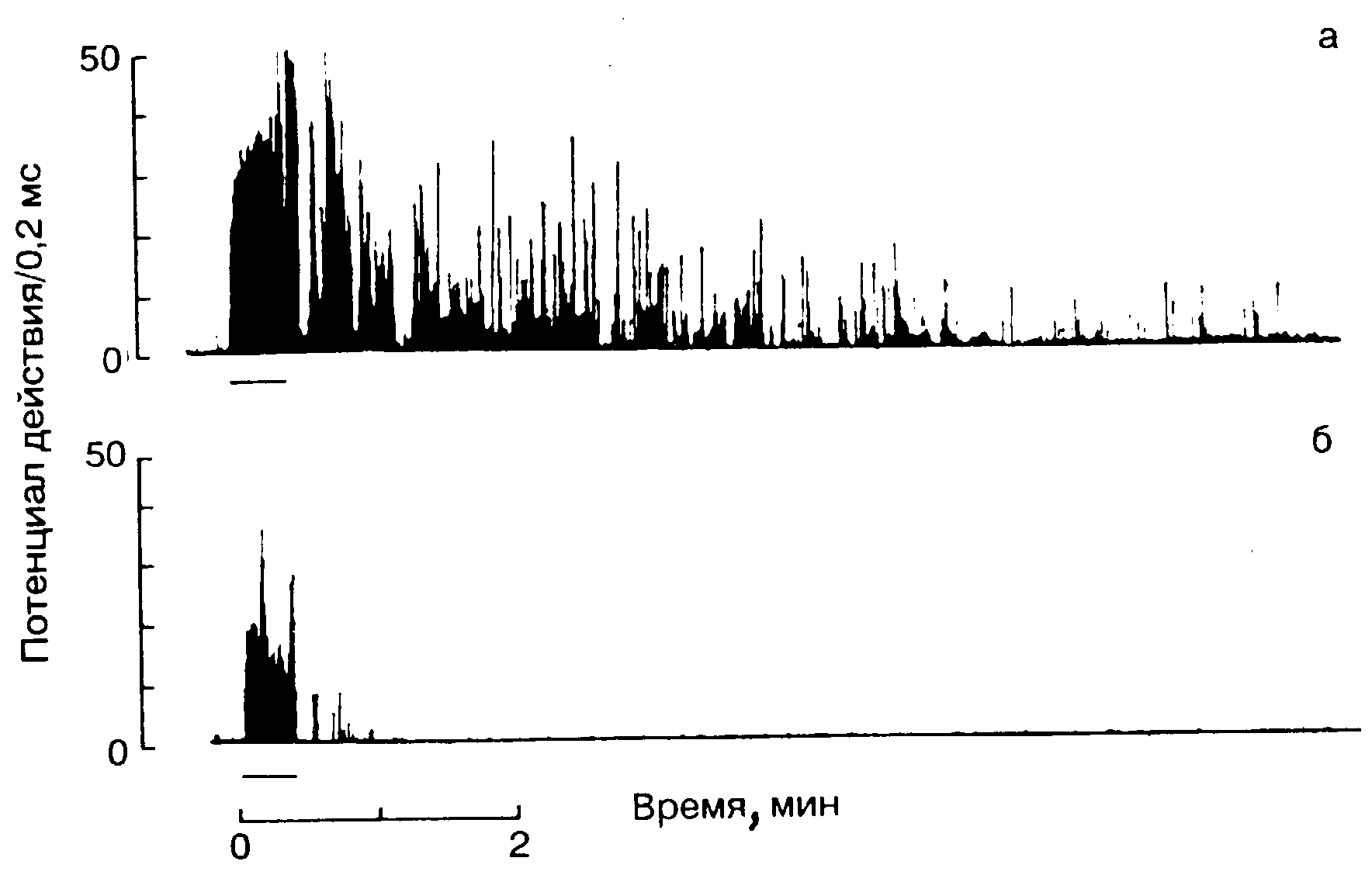
Рис. 12-6. Центральная гиперсенсибилизация при перевозбуждении.
Частота разрядов, возникающих в а-моторных нейронах заднего двуглавого нерва бедра у крысы при стимуляции хронически пересекаемого седалищного нерва 20 с, 10 Гц (на С-волокна). Период стимуляции отмечен горизонтальной линией под каждым обозначением. а-стимуляция хронически пересекаемого нерва сопровождается очень длительным послеразрядным периодом («выветривание»); б-стимуляция неповрежденного седалищного нерва вызывает значительно меньшее количество потенциалов действия в течение более короткого времени. (По Wall и Woolf [66], с разрешения.)