

Вопрос 15: Метаболизм-обмен веществ.

Катаболизм-распад,получение энергии Амфиболизм-промежуточный обмен Анаболизм-построение, затрата энергии
Катаболизм-распад пит.веществ на небольшие фрагменты, которые в ходе реакций амфиболизма превращаются в ряд орг.кислот и фосфорных эфиров. Анаболизм-синтез более сложных в-тв (полимеров: белки, полисахариды, полинуклеотиды) из более простых (мономеры: аминокислоты, сахара, нуклеотиды).
В самом общем виде процессы, способные служить источником энергии для прокариот, можно представить следующим образом:
А = >В + е– .
Например,
Fe2+ = >Fe3+ + е– ; (1)
-CH2-CH2- => -CH2=CH2- + 2e– ; (2)
СН4 + 1/2 O2=> СН3ОН . (3)
В первой реакции окисление иона двухвалентного железа — это потеря электрона. Во втором примере окисление углеродного субстрата можно в равной мере рассматривать как отрыв от него водорода (дегидрирование) или независимое удаление двух протонов (Н+) и электронов (e–).В последнем примере имеет место присоединение атома кислорода к молекуле субстрата. Окислительно-восстановительный характер реакции в этом случае не столь очевиден, как в предыдущих, поскольку не происходит отрыва электрона (водорода) от молекулы метана. В данном случае в результате окисления метана происходит замена связи C-H на связь C-OH, кислород оттягивает электроны от атома углерода, подвергшегося окислению, сам при этом восстанавливаясь. Таким образом, внутри молекулы происходит "деление" электронной пары между атомами углерода и кислорода, т. е. окислительно-восстановительные перестройки.
Т.о., реакции, в которых имеется возможность отрыва электронов, могут быть использованы прокариотами для получения энергии.
У прокариот известны три способа получения энергии:
1) разные виды брожения(СУБСТРАТНОЕ ФОСФОРИЛИРОВАНИЕ)
2) дыхание (ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ)
3)фотосинтез.
1) В процессах брожения в определенных окислительно-восстановительных реакциях образуются нестабильные молекулы, фосфатная группа которых содержит много свободной энергии. Эта группа с помощью соответствующего фермента переносится на молекулу АДФ, что приводит к образованию АТФ. Реакции, в которых энергия, освобождающаяся на определенных окислительных этапах брожения запасается в молекулах АТФ, получили название субстратного фосфорилирования.
2) При дыхании микроорганизмы(мко) окисляют восстановленные вещества с относительно низким окислительно-восстановительным потенциалом (E0), возникающие в реакциях промежуточного метаболизма или являющиеся исходными субстратами, например НАД·H2, сукцинат, лактат, NH3,
Окисление идет в результате переноса электронов через электронтранспортную цепь (ЭТЦ) .Окисляться могут как органические, так и неорг. соединения (H2, NH4+, NO2–, H2S, S0, S2O32–, Fe2+ и др).
3
Зависимый от бактериохлорофилла, бескислородный (зеленый,пурпурные бактерии)
Зависимый от бактериородопсина, бескислородный (галофильные архебактерии). Без окислительно-восстановительных переносчиков!.В этом случае энергия возникает в результате светозависимого перемещения протонов через мембрану!
Зависимый от хлорофилла, кислородный (цианобактерии)
)фотосинтез
В процессах дыхания и фотосинтеза освобождающаяся при переносе электронов энергия запасается первоначально в форме электрохимического трансмембранного градиента ионов водорода , которая затем может быть использована для синтеза АТФ.
Энергетическая валюта


МАКРОЭРГИ (АТФ, ФОСФОЕНОЛПИРУВАТ-ФЕП, ГЛЮКОЗА-1 ФОСФАТ)
МЕМБРАННЫЙ ПРОТОННЫЙ ИЛИ НАТРИЕВЫЙ ПОТЕНЦИАЛ
Химическая(энергия высокоэнерг.соединений)
Электрохимическая(энергия трансмембранного потенциала)
Центральное место в процессах переноса химической энергии принадлежит системе АТФ. АТФ образуется в реакциях субстратного и мембранзависимого фосфорилирования.
Преимущества АТР:
1)При гидролизе АТР обр-ся свободная энергия=-32кДж/моль
2)химически АТР высокостабильна (не идет без участия ферментов)
3) по об-ию свободной энергии: ФЕП >АТР>глюкоза-1-фосфат
-58кДж > -32кДж > -9кДж
Поэтому АТР может переносить энергию от сверхэнергетических соединений к низкоэнерг.
Протонный градиент
H+ — химическая частица, несущая положительный заряд, неравномерное его накопление по обе стороны мембраны приводит к возникновению ориентированного поперек мембраны электрического поля (суммарный положительный заряд, где происходит накопление H+, и отрицательный заряд по другую сторону мембраны).
Расположение переносчиков электронов в ЦПМ прокариот таково, что при работе любой электронтранспортной цепи (фотосинтетической или дыхательной) во внешней среде происходит накопление ионов водорода (протонов), приводящее к подкислению среды, а в клеточной цитоплазме — их уменьшение, сопровождающееся ее подщелочением, т. е. на мембране возникает ориентированный поперек (трансмембранный) градиент ионов водорода.
Итак, в соответствии с хемиосмотической теорией П. Митчелла, энергия, освобождаемая в результате работы; электронтранспортной цепи, первоначально накапливается в форме трансмембранного градиента ионов водорода. Разрядка образующегося DmH+ происходит с участием локализованного в той же мембране протонного АТФ-синтазного комплекса: Н+ возвращаются по градиенту DmH+ через Н+–АТФ-синтазу, при этом из АДФ и неорганического фосфата образуется АТФ. Предположительно, для синтеза одной молекулы АТФ достаточен перенос двух протонов, т. е. Н+/АТФ=2.
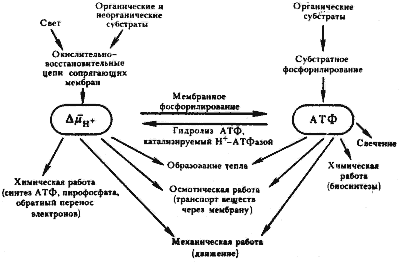
Преобразование энергии в клетке прокариот (по Скулачеву, 1980)
Натриевый градиент галофильных бактерий.
Галофильные бактерии обитают в экстремальных условиях среды (избыток солей NaCl).За счет света с помощью бактериродопсина они вырабатывают энергию в виде протонного градиента. Такая энергия хватает только днем. А ночью? Ночью они получают энергию за счет работы K-Na градиента.
Такой источник энергии складывается из разности хим=х потенциалов натрия по 2 стороны ЦПМ и разности жлектрического потенциалов на противоположных поверхностях мембраны.
Натриевый градиент обеспечивает энергией: процесс фосфорилирования АДР; активный транспорт, вращение жгутиков и т.д.
Законы биоэнергетики
1) Живая клетка избегает прямого использования энергии внешних ресурсов для совершения полезной работы. Она сначала превращает их в одну из трех конвертируемых форм энергии ("энергетических валют")
2) Любая живая клетка всегда располагает как минимум двумя "энергетическими валютами": водорастворимой (АТФ ) и связанной с мембраной
3) "Энергетические валюты" клетки могут превращаться одна в другую. Поэтому получения хотя бы одной из них за счет внешних ресурсов достаточно для поддержания жизнедеятельности.
16вопрос.=)
ГИПОМЕТАБОЛИЗМ-снижение обмена веществ, позволяющее сократить энерг.затраты в небл.условиях.
Цистоподобные рефрактерные клетки
Специализированные анабиотические формы (такие как эндо- и экзоспоры, миксоспоры, цисты) известны лишь для ограниченного круга микроорганизмов. Однако как у спорообразующих микроорганизмов (Bacillus cereus) на фоне подавления спорообразования, так и у споронеобразующих (Pseudomonas carboxidoflava, Micrococcus luteus, Escherichia coli) бактерий внеклеточные микробные метаболиты, проявляющие свойства индукторов анабиоза, вызывают образование цистоподобных рефрактерных клеток (термин связан с переходом целой клетки в покоящуюся форму и увеличенной способностью данных клеток по сравнению с обычными сильно преломлять свет).
В этих клетках снижение метаболической активности (гипометаболизм) сопровождается развитием резистентности к экстремальным воздействиям. Эти формы отличаются нерегистрируемым уровнем эндогенного дыхания, повышенной терморезистентностью и специфической ультраструктурой. Для споронеобразующих микроорганизмов цистоподобные рефрактерные клетки — единственная покоящаяся форма, способная сохраняться в течение нескольких лет, а для спорообразующих бактерий — альтернатива спорообразования.
Некультивируемые формы
Главная особенность этого состояния заключается в том, что такие бактерии не размножаются и поэтому не образуют колоний на плотной питательной среде. Такие не размножающиеся, но жизнеспособные клетки получили название некультивируемых форм бактерий (НФБ). Клетки НФБ, находящиеся в некультивируемом состоянии (НС), обладают активными метаболическими системами, в том числе системами переноса электронов, биосинтеза белка и нуклеиновых кислот, и сохраняют вирулентность. Их клеточная мембрана более вязкая, клетки обычно приобретают форму кокков, имеют значительно уменьшенные размеры. НФБ обладают более высокой устойчивостью во внешней среде и поэтому могут переживать в ней длительное время (например, холерный вибрион в грязном водоеме), поддерживая эндемическое состояние данного региона (водоема).
Для обнаружения НФБ используют молекулярно-генетические методы (ДНК—ДНК-гибридизация, ЦПР), а также более простой метод прямого подсчета жизнеспособных клеток. С этой целью к исследуемому материалу добавляют в небольшом количестве питательные вещества (дрожжевой экстракт) и налидиксовую кислоту (для подавления синтеза ДНК) на несколько часов. Клетки усваивают питательные вещества и увеличиваются в размерах, но не делятся, поэтому такие увеличенные клетки четко видны в микроскоп и их легко подсчитать. Генетические механизмы, обусловливающие переход бактерий в НС и их реверсию из него, не ясны.
