

1 Вопрос
Рост прокариотной клетки ‐ согласованное увеличение количества всех химических
компонентов, из которых она построена. Рост приводит к увеличению массы (а
следовательно, и размеров) клетки.
Рост клетки не беспределен. После достижения определенных (критических) размеров
клетка подвергается размножению – увеличение числа клеток микроорганизмов в
популяции. Кроме состава питательной среды, рост бактерий сильно зависит от условий окружающей среды, поэтому эти условия нужно строго контролировать. Микроорганизмы лучше растут при определенных температурах и рН, и они чувствительны к ряду факторов, таких как концентрация кислорода и интенсивность света. Оптимальными являются условия, при которых обеспечивается самое короткое время генерации. Каждый вид микроорганизмов приспосабливается к определенным условиям окружающей среды. Под удельной скоростью роста понимают часовой прирост, пересчи-
танный на единицу растущей массы. Продолжительность жизни одного поколения (время генерации).между временем генерации и удельной скоростью роста су-
ществует обратно пропорциональная зависимость. Скорость роста не явля-
ется величиной неизменной. Константу скорости деления, или число клеточных делений в единицу времени, t−t0 можно вычислить по формуле: ν = n/t,
а время одной генерации (g), по формуле: g = t/n = 1/ν. Прирост биомасс называется урожаем. Урожай клеток отражает интенсивность развития культуры. Под урожаем клеток (Х) понимают разность между максимальной и исходной массой бактерий: X = Xmaх – Х0. Эту величину выражают в весовых единицах, чаще в граммах.
Периодическое культивирование.выращивание микроорганизмов на несменяемой среде от инокуляции до окончания роста клеток вследствие исчерпания питательных субстратов или накопления вредных веществ. В отличие от периодического культивирования в непрерывных процессах питательная среда подается непрерывно, удаление биомассы и продуктов ее жизнедеятельности также осуществляется непрерывно.
Установившиеся режимы непрерывного культивирования характеризуются постоянством концентрации микроорганизмов и удельной скорости роста популяции. Длительного удержания клеток в системе (диализная культура). В этом случае продлевается экспоненциальная фаза и фаза линейного роста. Суть процесса диализ заключается в том, что культура развивается в пространстве, ограниченном полупроницаемой мембраной, а продукты метаболизма диффундируют во внешний раствор. Наиболее простой диализный метод – культивирование в целлофановых мешках, погруженных в питательную среду.
1. Начальная фаза. промежуток времени между инокуляцией и
достижением максимальной скорости деления. Это период адаптации бактериальной популяции.
Продолжительность этой фазы зависит от:
‐ предшествовавших условий культивирования и возраста инокулята,
‐ от того, насколько пригодна для роста данная среда (количества и качества питательной
среды).
2. Экспоненциальная (логарифмическая) фаза. Численность бактериальной популяции
увеличивается в геометрической прогрессии. Характеризуется постоянной максимальной
скоростью деления клеток. Эта скорость зависит от вида бактерий и от среды.3. Стационарная фаза. Стационарная фаза наступает тогда, когда число клеток перестает увеличиваться. Т.е. наступает равновесие между количеством погибших, вновь образующихся и находящихся в состоянии покоя клеток.Скорость роста снижается, время генерации – увеличивается. Это происходит в результате истощения питательной среды, накопления в ней продуктов метаболизма и дефицита кислорода. Количество биомассы, достигнутое в стационарной фазе, называют выходом или урожаем. Урожай зависит от природы и количества используемых питательных веществ, а также от условий культивирования. 4. Фаза отмирания. Происходит гибель бактериальных клеток. Число живых клеток может снижаться экспоненциально. Сравнительно легко понять случаи, когда в среде накапливаются кислоты (при росте Escherichia, Lactobacillus). Причины гибели в др. случаях более трудно объяснимы.
2 вопрос.Отношение микроорганизмов к молекулярному кислороду. Кислород как акцептор электронов в дыхательной цепи. Ферменты детоксикации активных видов кислорода: каталаза, супероксиддисмутаза. Кислород как окислитель. Моно- и диоксигеназы.
Кислород широко распространен в природе и находится в связанном (вода, орг. И неорг. Соед.) и индивидуальном состоянии – в атмосфере в виде молекулярного кислорода.
Объемная доля молекулярного кислорода – 21%.
Большинство мко удовлетворяет свои потребности в нем, используя обе формы кислорода.
Аэробы |
Анаэробы |
||||
облигатные |
факультативные |
облигатные |
|||
Растут при содержании O2 как на воздухе |
микроаэрофилы |
капнофилы |
аэротолерантные |
«строгие» |
|
|
растут при пониженном парциальном давлении О2 (2%) |
растут при пониженном парциальном давлении О2 и повышенном парциальном давлении СО2 (10-15%) |
способны расти в присутствии атмосферного кислорода, но не использовать его в качестве источника энергии, имеют ферменты, обезвреживающие реактивные формы О2 |
Растут в бескислородной среде, для них О2 - яд |
|
Большинство мко |
Helicobacter pylori, Streptococcus intermedius, азотфиксаторы |
Brucella abortus |
Маслянокислые, молочнокислые бактерии |
Bacteroides , Fusobacterium |
|
1: Облигатно аэробные (нуждающиеся в кислороде) бактерии собираются в верхней части пробирки, чтобы поглощать максимальное количество кислорода. (Исключение: микобактерии — рост пленкой на поверхности из-за восколипидной мембраны.)
2: Облигатные анаэробные бактерии собираются в нижней части, избегая кислорода (либо не дают роста).
3: Факультативно аэробные бактерии собираются в основном в верхней части, поскольку аэробное дыхание наиболее эффективно, однако они могут быть найдены на всем протяжении среды, так как недостаток кислорода не причиняет им вреда.
4: Микроаэрофилы собираются в верхней части пробирки, но не у самой поверхности, так как им требуется низкая концентрация кислорода.
5: Аэротолерантные анаэробы не реагируют на концентрацию кислорода и равномерно распределяются по пробирке.
Кислород служит конечным акцептором электронов при аэробном дыхании и поэтому нужен всем аэробам.
О2+4е=О2-+О2-
Реакцию катализирует цитохромоксидаза – конечный фермент ЭТЦ. Здесь одновременно происходит перенос 4е, и в результате получаются 2 иона О2-, каждый из которых при взаимодействии с 2Н+ образует воду.
У всех облигатных и факультативных аэробов.
O2+2e=O22- Реакция характерна для некоторых ферментов, содержащих флавин (глюкозооксидаза, оксидазы аминокислот). Эти ферменты одновременно переносят 2е и восстанавливают О2 до иона пероксида O22-, который, реагируя с протонами образует пероксид водорода. Перекись токсична, т.к окисляет, напр, SH-группы белков, повреждает ДНК. Защитное действие оказывают каталаза и пероксидаза.
Перекись+ каталаза=вода и кислород (донор е – перекись).
О2+1е=О2- Реакция катализируется многами оксидазами(NADPH-оксидаза), здесь переносится только 1е. при этом образуется супероксид-ион – это радикал, очень реакционноспособен. Защитное действие оказывает супероксиддисмутаза:
2О2-+2Н+ супероксиддисмутаза= пероксид+ кисолород.
Совместно с каталазой она превращает радикалы в безвредный кислород.
Хемотрофы(E.Coli), Аэротолерантные анаэробы, облигатные анаэробы(Clostridium).
Кислород как окислитель
1.Неферментативное окисление (поглощение кислорода суспензией убитых клеток, окисление хим. Веществ в клетке,напр, SH-групп белков.)
2.Ферментативное поглощение кислорода (дыхание):
Свободное окисление (энергия не запасается в выделяется в виде тепла). Сюда относятся реакции катализируемые моно- и диоксигеназами.
Окисление, сопряженное с запасанием Е (кислород - конечный акцептор электронов):
Не связанное с фосфорилированием(т.е окисление не связано с трансформированием свободной энергии в форму макроэргических фосфорных связей(АТФ). Тут образуются метаболиты, содержащие богатые энергией связи( например тиоэфирные C-S-KoA).
Фосфорилирующее окисление(Е высвобождаемая при электронном транспорте на молекулярный кислород и возникающая сначала в форме протонного потенциала, с помощью протонной АТФ-синтазы превращается в АТФ.
По химическим механизмам, все реакции этой классификации можно разделить на 2 типа.
Реакции, катализируемые кислородными трансферазами – диоксидазами.
Прямое присоединение молекулярного кислорода к молекуле метаболита: А+О2=АО2
Акцепторами кислорода могут быть молекулы 2 разных субстратов: Х+У+О2=ХО+УО.
Все такие реакции –свободное окисление и не связаны с получением Е.
Электроны идут к кислороду, выполняющему функцию конечного акцептора.
В этом случае 1, 2 или 4 е связываются с О2 и приводят его к неполному или полному восстановлению. Реакции катализируются оксидазами и могут быть реакциями свободного окисления или связанные с запасанием энергии.
Промежуточные реакции – судьба каждого из 2х атомов кислорода различна:
А+О2+ДН2=АО+Н2О+Д. АО-прямое присоединение, Н2О полное восстановление.
Обе реакции катализируются одним ферментом – монооксидазой со смешанными функциями. Запасание энергии маловероятно.
3 вопрос. Морфология микроорганизмов. Основные морфотипы микробных клеток. Типы агрегации клеток микроорганизмов. Внешние структуры микробной клетки. Слизистые слои, капсулы, чехлы, газовые баллоны, шипы, трубчатые выросты, целлюлосомы.
Основные морфотипы микробных клеток.
П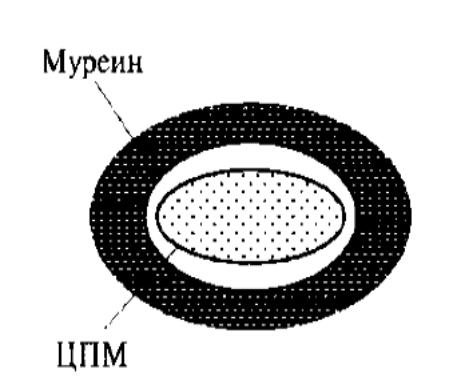 о
строению клеточной стенки.
Грамположительный
морфотип:20-40 слоев муреина (>10% сухой
массы клетки) плотно прилегают к ЦПМ,
нет наружной мембраны, тейхоевые кислоты
поперечно пронизывают весь саккулус
(муреиновый мешок) и выходя на поверхность
являются основными антигенами грам+
бактерий. Пептидные хвосты муриена
связаны перекрестносвязывающими
мостиками. Липидов мало.
о
строению клеточной стенки.
Грамположительный
морфотип:20-40 слоев муреина (>10% сухой
массы клетки) плотно прилегают к ЦПМ,
нет наружной мембраны, тейхоевые кислоты
поперечно пронизывают весь саккулус
(муреиновый мешок) и выходя на поверхность
являются основными антигенами грам+
бактерий. Пептидные хвосты муриена
связаны перекрестносвязывающими
мостиками. Липидов мало.
Г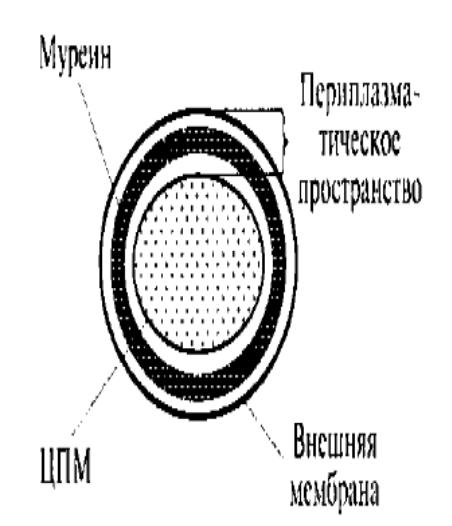 рамотрицательный
морфотип: 1-3 слоя муреина (<10% сухого
вещества клетки) неплотно прилегает к
мембране, есть наружная мембрана, между
наружной мембраной и ЦПМ – периплазматическое
пространство со слоем муреина внутри
него. Наружная мембрана содержит
фосфолипиды на внутренней поверхности
и липополисахариды – на внешней. Нет
тейхоевых кислот. Пептидные хвосты
муреина не связываются, а если связываются,
то напрямую, без перекрест. мостиков.
Липидов много, 80% сухой массы клеточной
стенки. Муреин связан с наружной мембраной
липопротеидами Брауна.
рамотрицательный
морфотип: 1-3 слоя муреина (<10% сухого
вещества клетки) неплотно прилегает к
мембране, есть наружная мембрана, между
наружной мембраной и ЦПМ – периплазматическое
пространство со слоем муреина внутри
него. Наружная мембрана содержит
фосфолипиды на внутренней поверхности
и липополисахариды – на внешней. Нет
тейхоевых кислот. Пептидные хвосты
муреина не связываются, а если связываются,
то напрямую, без перекрест. мостиков.
Липидов много, 80% сухой массы клеточной
стенки. Муреин связан с наружной мембраной
липопротеидами Брауна.
Типы агрегации клеток микроорганизмов.
Диплококки – пары кокков (деление в 1 плоскости).
Диплобациллы – пары палочек (деление в 1 плоскости).
Стрептококки – короткие и длинные цепочки кокков (деление в 1 плоскости).
Стрептобациллы – короткие и длинные цепочки палочек (деление в 1 плоскости).
Тетракокки – 4 кокка (деление в 2 перпендикулярных плоскостях). Сарцины – пакеты из 8, 12, 16 кокков (деление в 3 перпендикулярных плоскостях).
Стафилококки – гроздья винограда (деление в 2 перпендикулярных плоскостях).
Внешние структуры микробной клетки.
Внешние структуры – структуры, расположенные снаружи от ЦПМ: жгутики, ворсинки, капсула, чехол, слизистый слой, S-слой, клеточная стенка, газовые баллоны, целлюлосомы. Иногда ЦПМ также относят к поверхностным структурам. ЦПМ – единственная обязательная поверхностная структура. Остальные необязательны, но создают адаптационные преимущества.
К леточная
стенка+капсула/чехол/слизь/S-слой=клеточная
оболочка.
леточная
стенка+капсула/чехол/слизь/S-слой=клеточная
оболочка.
Слизистые слои, капсулы, чехлы, газовые баллоны, шипы, трубчатые выросты, целлюлосомы.
1.Покровы. Их могут образовывать как отдельные клетки, так и группы клеток. Состоят изз полисахаридов, белков, липидов, фосфатов.
Слизистые слои – имеют аморфный, бесструктурный вид. Неплотно связана с клеточной стенкой. Имеют однородное строение. Образуются снаружи от клетки из сахаров окружающей среды за счет выделяемых клеткой полимеризующих ферментов. Leuconostoc mesenteroides – вредитель сахарной свеклы, используется в молочной промышленности. Сахарозу полимеризует в глюканы, образуя слизь, поэтому свекла портится там, где много сахарозы. Streptococcus mutans, Streptococcus salivarius – во рту, полимеризуют сахарозу до леванов, образуют молочную кислоту, а их слизь – матрикс для молочной кислоты, а она разрушает зубную эмаль, что ведет к кариесу.
Капсула – имеет четко очерченные внешние границы, плотно связана с клеточной стенкой, имеет однородное строение (из полисахаридов и 99% воды). Фактор вирулентности: сама антиген и защищает от антибиотиков. Наличие ее зависит от штамма и условий культивирования (в лабораторных условиях частично теряется). Если гладкие S-колонии – капсула имеется, если шероховатые R-колонии – капсулы нет. Классификация:
I. Микрокапсулы (толщина <0,2 мкм) и макрокапсулы (>0,2 мкм).
II. 1. из гомо- и гетерополисахаридов: альгинаты, декстран, ксантан, леван, целлюлоза. Leuconostoc mesenteroides (из декстрана), Acetobacter xylinum (из целлюлозы).
2. из полипептида. Bacillus anthracis (из поли-D-глутамата).
3. из полифосфата. Neisseria meningitidis (из конденсированного неорганического фосфата).
Чехлы (влагалища) – имеют четко очерченные внешние границы, плотно связаны с клеточной стенкой, имеют тонкую структуру, состоят из сахаров, гексозаминов, белков, липидов, фосфатов, мб инкрустированы окислами Mg, Fe, участвуют в биоминерализации. Их образуют вегетативные клетки (хламидобактерии Sphaerotilus natans, Leptothrix ocharacea), цианобактерии (одноклеточные и трихомные) и дифференцированные клетки (эндоспоры, гетероцисты цианобактерий). У хламидобактерий – полая трубка с 1-3 цепочками клеток внутри.
2. Поверхностные органеллы. Шипы (спины) – жесткий полый цилиндр из белка спинина, уложенного по спирали. Перпендикулярный клетке. Прикреплены с наружной поверхности наружной мембраны, но не проникают, в месте прикрепления могут расширяться (воронка). Около 10 шипов на клетку. Долговечны, прочны, устойчивы к сильным кислотам и щелочам, их функция неизвестна, вероятно, обеспечивают лучшее выживание (на их синтез идет много белка, не просто так). Обеспечивают плавучесть, агрегацию, контакт между клетками, обмен генетической информацией, защищают от фаготрофных процистов, мб для адаптации к солености и температуре, т.к. образование шипов индуцируется этими факторами. У грамотрицательных (псевдомонады, метилотрофы, цианобактерии).
Экстрацеллюлярные газовые баллоны – крупные газонакопительные везикулы сферической, цилиндрической или игольчатой формы, сформированы неунитарной мембраной, прикреплены к наружной поверхности клетки. Встречаются редко, Bacillus vesiculi fereus.
Целлюлосома – самая крупная поверхностная органелла сферической формы, функции – биодеградация целлюлозы, обеспечение лучшей адгезии к субстрату, лучшего взаимодействия целлюлаз с субстратом, канализированное поступление продуктов гидролиза к поверхности клетки.
Пили (фимбрии, ворсинки) – прямые белковые цилиндры из белка пилина (фимбрина), субъединицы уложены по спирали. 1-тысячи на клетку. Главная функция – адгезия клеток к субстрату или к др бактериям. Расположены полярно (F-пили) или перитрихиально.
а) пили I типа – адгезия, обеспечивают гидрофобность, являются фактором патогенности и вирулентности, т.к. вызывают агглютинацию эритроцитов. При выращивании клеток на твердой среде пили не образуются, а в жидкой среде без аэрации пили образуются, формируя биопленки. Их формирование кодируется хромосомными генами.
б) пили IV типа – могут сокращаться, обеспечивая движение рывком (твитчинг)
в) половые пили (F-пили) – их формирование кодируется плазмидными генами. Эта плазмида – фактор трансмиссивности. Клетки, несущие ее – доноры, а не несущие – реципиенты. Половые пили обеспечивают взаимную адгезию бактерий при образовании конъюгативных пар.
4 вопрос. Движение бактерий. Структуры, обеспечивающие различные типы движения бактерий. Жгутики, фимбрии. Молекулярное строение жгутикового аппарата про- и эукариотных микроорганизмов. Понятия хемо- и фототаксиса.
10% всех бактерий способны к движению. Это адаптационное преимущество. Существуют разные механизмы движения. 1. С помощью жгутиков:
а) плавание – по жидкой среде.
б) с помощью периплазматических эндожгутиков (аксисальных нитей). Аксиальная нить – комплекс жгутиков, тянущихся от одного конца клетки к другому, по спирали оборачиваясь вокруг нее.
в) «роение» с помощью многочисленных жгутиков – по твердой среде. Осуществляют многожгутиковые клетки-швермеры, они формируют колонию и способствуют распространению популяции. Характерно для патогенных мко. (Proteus) 2. Подтягивание с помощью пилей IV типа. У многих грамм-. (Pseudomonas, Neisseria). Резкое укорочение и удлинение пилей. Образование биопленок.
3. Скольжение – с помощью слизи для смачивания твердой поверхности для улучшения скольжения. Механизм развит у всех.
4. Движение на хозяйском актине
Типы бактерий-жгутиконосцев (по числу и расположению жгутиков):
Монотрих – монополярное монотрихиальное жгутикование (Vibrio cholerae).
Лофотрих – монополярное политрихиальное (Pseudomonas, Thiospirillum)
Амфитрих – биполярное моно- или политрихиальное (Spirillum)
Перитрих – полиполярное политрихиальное (E. coli, Salmonella enterica)
Молекулярное строение жгутикового аппарата про- и эукариотных микроорганизмов.
Прокариоты. 3 основные структуры жгутика:
1)Филамент (фибрилла, пропеллер) — полая белковая нить из 11 рядов субъединиц флагеллина, уложенного по спирали. Полость внутри используется при синтезе жгутика — он происходит в направлении от ЦПМ. По полости к собираемому в настоящий момент участку переносятся субъединицы флагеллина.
2)Крюк — более толстое, чем филамент, белковое (не флагеллиновое) образование.
3)Базальное тело (трансмембранный мотор)
а) стержень
б)диски (кольца), нанизанные на стержень: М-диск (основной приводящий в движение жгутик), сверху – S-диск. У грам- они находятся в периплазматическом пространстве, а у грам+ - в пептидогликановом слое. Сейчас их считают единым MS-кольцом. Ещё два кольца — P и L — есть только у грамотрицательных бактерий, они расположены в пептидогликановом слое и наружной мембране соответственно, неподвижны и лишь направляют стержень ротора мотора. Вокруг MS-кольца в ЦПМ расположены статоры — белковые комплексы MotA4/MotB4 представляющие собой протонный канал.
4)Белковый колпачок, утопленный в цитозоле.
Точный механизм работы базального тела не известен. Большинство исследователей полагает что поступление протона из периплазмы или внешней среды в MotA4/MotB4 комплекс вызывает конформационные изменения белков, благодаря электростатическому взаимодействию или прямому контакту это изменение приводит к повороту MS-кольца, а его дальнейшее движение возвращает исходную конформацию комплексу и выталкивает протон в цитозоль. У Escherichia coli для одного оборота жгутика требуется перемещение около 1000 протонов. Таким образом, базальное тело преобразует химическую энергию в работу, вращаясь за счёт градиента концентрации протонов или, в редких случаях, ионов натрия (некоторые морские бактерии рода Vibrio, алкалофильные Bacillus, Acetobacterium woodii), это вращение осуществляется со скоростью до 100 об/сек, причём его направление может изменяться менее чем за 0,1 сек.
Средняя скорость движдения бактерии – 20-200 мкм/с.
М-диск вращается против часовой стрелки со скоростью 300 об/сек, жгутик – 40-60 об/сек, а сама бактерия – 12-14 об/мин по часовой стрелке (поступательное движение). Подавляющее большинство наделённых жгутиком бактерий имеют палочковидную форму. Из гидродинамических расчётов следует, что для наиболее эффективного движения отношение длины клетки к ширине должно составлять 3,7. Движение кокков крайне неэффективно, поэтому они чаще всего неподвижны.
Бактерии-перитрихи собирают при движении все свои жгутики (каждый из которых вращается против часовой стрелки) в один пучок. Толщина 10—20 нм и длина 3—15 мкм. При вращении жгутика по часовой стрелке клетка тянется за жгутиком (жгутик впереди), при вращении жгутика против часовой стрелки жгутик выталкивает клетку вперед (жгутик сзади). Перитрихи вращают пучок жгутиков против часовой стрелки (жгутики сзади). Если надо поменять направление движения, клетка останавливается и делает кувырок.
Эукариоты. Тонкий вырост на поверхности клетки, одетый трёхслойной клеточной мембраной и содержащий аксонему — совокупность микротрубочек из белка динеина (9 периферических дуплетов по окружности и 2 центральных микротрубочек-синглетов). Каждый дуплет состоит из полной А-микротрубочки и неполной В-микротрубочки. В-трубочка диплета соединяется с А-трубочкой следующего динеиновыми ручками. Базальное тело (кинетосома) – внутриклеточная часть жгутика.
Жгутик осуществляет движение со скоростью 10-40 об/сек. Толщина до 200 нм и длина до 100 мкм.
Понятия хемо- и фототаксиса.
Таксис – направленное движение у бактерий в ответ на внешний раздражитель, простейшая поведенческая реакция бактерий.
Фототаксис – движение мко в направлении или от источника света. Фототаксис называется положительным, если движение происходит по направлению к источнику света и отрицательным, если в противоположном направлении.
Хемотаксис – двигательная реакция микроорганизмов на химический раздражитель.
Эффекторы, вызывающие хемотаксис, делятся на аттрактанты (например, О2 для аэробов) и репелленты (например, О2 для анаэробов).
5 вопрос.Остов пептидогликана одинаков у всех бактерий и состоит из чередующихся молекул аминосахаров — N-ацетилглюкозамина (N-АцГлю) и N-ацетилмураминовой кислоты (N-АцМур), связанных между собой р-гликозидными связями. Боковые цепочки в каждой молекуле пептидогликана представлены набором идентичных тетрапептидов. Поперечные цепочки также представлены набором из идентичных для данной молекулы пептидогликана пентапептидов, содержащих глицин, — пентаглицинов, однако у разных видов бактерий боковые и поперечные пептиды различны.
В тетрапептидной боковой цепочке у большинства грамотрицательных бактерий имеется диаминопимелиновая (диаминопимеловая) кислота (ДАП) — уникальный компонент клеточной стенки, обнаруженный только у прокариот. Кроме того, в составе боковых цепочек пептидогликана обнаружены D-аминокислоты (D-аланин, D-глутамин). Боковые тетрапептиды связаны с N-ацетилмураминовой кислотой остова. Связывание боковых тетрапептидов между собой происходит путем образования поперечных пентаглициновых мостиков между D-аланином одной цепи и диаминопимелиновой кислотой (или иной аминокислотой) другого бокового пептида.
Наличие двух типов связей (гликозидные и пептидные), которые соединяют субъединицы пептидогликанов, придает этому гетерополимеру структуру молекулярной сети. Благодаря этим связям пептидогликановый слой клеточной стенки образует огромного размера ригидную мешковидную макромолекулу, которая окружает протопласт, уравновешивает его тургорное давление (у Е. coli — до 15 атм.) и придает ему определенную постоянную форму. Пептидогликан может разрушаться под действием различных ферментов, а его синтез блокируют бета-лактамные антибиотики.
Связь между N-ацетилмураминовой кислотой и N-ацетилглюкозамином разрушается лизоцимом, связь между N-ацетилмураминовой кислотой и боковым пептидом (его L-аланином) расщепляют амидазы, а связи межпептидные — эндопептидазы. Пентаглициновый мостик стафилококкового пептидогликана разрушается лизоста-фином. Образование поперечных сшивок между боковыми цепочками тетрапептидов блокируется пенициллинами (беталактамными антибиотиками). Это приводит к разрыхлению пептидогликановой сети, следствием чего является осмотический лизис растущих клеток. Пептидогликан, помимо того что он определяет постоянную форму бактерий, обладает следующими важнейшими иммунобиологическими свойствами.
1. В его составе обнаружены родоспецифические антигенные детерминанты. Они содержатся в гликановом остове и в тетрапептидах. В межпептидных мостиках имеются видоспецифические антигенные детерминанты.
2. Пептидогликан запускает классический и альтернативный пути активации системы комплемента.
3. Он тормозит фагоцитарную активность макрофагов, т. е. защищает бактерии, особенно грамположительные, от фагоцитоза.
4. Угнетает миграцию макрофагов.
5. Способен индуцировать развитие гиперчувствительности замедленного действия.
6. Обладает противоопухолевым действием.
7. Оказывает пирогенное действие на организм человека и животных.
Таким образом, клеточная стенка является чрезвычайно важной биологической структурой бактерий, определяющей многие их специфические свойства. Как отмечалось выше, все бактерии, в зависимости от их отношения к окраске по Граму, делятся на грамположительные и грамотрицательные. Суть окраски по Граму заключается в том, что вначале бактерии окрашивают кристаллическим или генциановым фиолетовым, а затем — раствором Люголя, после чего мазок обрабатывают спиртом и докрашивают водным фуксином. Грамотрицательные бактерии обесцвечиваются спиртом и поэтому окрашиваются в красный цвет, а грамположительные не обесцвечиваются и сохраняют фиолетовую окраску. Это свойство грамположительных бактерий зависит исключительно от особенностей химического состава и структуры их клеточных стенок, так как при разрушении клеточных стенок или утрате их (в случае L-трансформации) они становятся грамотрицательными.
Причину различного отношения бактерий к окраске по Граму объясняют тем, что после обработки раствором Люголя образуется не растворимый в спирте комплекс йода с генциановым фиолетовым, который у грамположительных бактерий в связи со слабой проницаемостью их стенки не может диффундировать из клетки, в то время как у грам-отрицательных легко удаляется при промывании их этанолом, а затем водой.Большинство архей (но не Thermoplasma и Ferroplasma) обладают клеточной стенкой]. У большей части из них она сформирована молекулами поверхностных белков, образующих наружный S-слой[]. S-слой представляет собой жёсткую сетку из белковых молекул, покрывающих клетку снаружи, подобно кольчуге[]. Этот слой защищает клетку от физических и химических воздействий, а также предотвращает контакт макромолекул с клеточной мембраной[. В отличие от бактерий, клеточная стенка архей не содержит пептидогликан[]. Метанобактерии (лат. Methanobacteriales) имеют клеточные стенки, содержащие псевдопептидогликан, который напоминает пептидогликан эубактерий по морфологии, функции и физической структуре, но отличен по химической: в нём нет остатков D-аминокислот и N-ацетилмурамовой кислоты Снаружи от клеточной мембраны большинство бактерий одето клеточной стенкой, несколько напоминающей целлюлозную стенку растительных клеток, но состоящей из других полимеров (в их состав входят не только углеводы, но и аминокислоты и специфические для бактерий вещества). Эта оболочка не дает бактериальной клетке лопнуть, когда в нее за счет осмоса поступает вода. Поверх клеточной стенки часто находится защитная слизистая капсула. Окраска по граму-метод дифференциальной окраски клеток прокариот, разработанный датским микробиол. X. Громом в 1884 г. Заключается в последовательной обработке мазка клеток основными красителями (напр., кристаллическим фиолетовым) и иодом и последующем отмывании его в этаноле или ацетоне. В зависимости от свойств клеточной стенки бактерий красители могут удерживаться ими в разной степени. Бактерии, сохраняющие окраску после обработки растворителями, получили название грамположительных (Гр+), а обесцвечивающиеся – грамотрицательных (Гр–). О. б. Г. – систематический признак.бактериальные S-слои (англ. bacterial S-Layers) — поверхностные структуры бактериальных клеток, состоящие из отдельных молекул белков или гликопротеидов, собранных в упорядоченный 2-мерный слой, который полностью покрывает поверхность клетки.
Толщина бактериального S-слоя находится в диапазоне 5–15 нм. Такой слой имеет поры, величина диаметра которых равномерно распределена в диапазоне 2–6 нм. Эти слои можно использовать в качестве компонентов наноэлектронных устройств и биосенсоров.ПРОТОПЛА́СТ (от прото... и греч. plastos — вылепленный, образованный), все содержимое клетки, за исключением клеточной оболочки.сферопласт
(сферо- + греч. plastos созданный, образованный) бактериальная клетка с частично разрушенной (редуцированной) клеточной стенкой, характеризующаяся неустойчивостью к изменениям осмотического давления.L-формы — бактерии, частично или полностью лишённые клеточной стенки, но сохранившие способность к развитию. Впервые обнаружены в 1894 году Н.Ф. Гамалеем.Было показано, что L-формы возникают спонтанно или индуцировано - под воздействием агентов, блокирующих синтез клеточной стенки: антибиотиков (пенициллины циклосерин, цефалоспорины, ванкомицин), ферментов (лизоцим, амидаза, эндопептидаза) ультрафиолетовых и рентгеновских лучей, аминокислоты глицина.Все L-формы независимо от вида бактерий, из которого они возникли, обладают общими особенностями:
Сходство морфологических изменений: образование нитевидных, волокнистых, колбасовидных, шаровидных и гранулярных форм.
Сходные культуральные свойства: анаэробные или микроаэрофильные условия роста, потребность в холестерине и сывороточном белке, рост на плотных средах в виде колоний двух типов А и В Колонии типа А растут на поверхности агара, имеют очень мелкие размеры. Они состоят главным образом из гранулярных структур, лишённых клеточной стенки, и очень похожи на микоплазмы. Колонии типа В состоят из центральной зоны, врастающей в агар, и прозрачной фестончатой периферической зоны. Они похожи по внешнему виду на колонии типа "глазуньи", образуемые микоплазмами, но более крупные и грубые. В этих колониях обнаруживаются крупные тела, содержащие компоненты клеточной стенки, сходные со стенкой родительских бактерий, но лишённые ригидности. Многие бактерии могут образовывать колонии А и В типов, однако грамположительные бактерии чаще образуют колонии только типа А L-формы из колоний типа В легко ревертируют в исходные формы. Колонии типа А более стабильны и ревертируют в исходные формы значительно реже.
Постепенное (по мере нарушения синтеза клеточной стенки) превращение из грамположительных в грамотрицательные структуры.
Образование стабильных и нестабильных L-форм (в зависимости от степени полноты утраты способности синтезировать клеточную стенку.)
Изменение антигенных свойств (утрата К- и О-антигенов, как следствие нарушения синтеза клеточной стенки).
Снижение вирулентности по сравнению с исходными родительскими формами в связи с утратой различных факторов патогенности (адгезии, инвазии, эндотоксина и т.п)
Способность длительно персистировать (переживать) в организме. Утрата клеточной стенки делает L-формы нечувствительными к различным химиопрепаратам и антителам.
Способности при неполной утрате синтеза клеточной стенки возвращаться в исходную бактериальную форму.R-форма
термин, используемый для обозначения бактерий, образующих на плотных питательных средах колонии неправильно-округлой формы, с зазубренными краями, плоской и морщинистой поверхностью
6 вопрос.Систематика-теория многообразия живых микроорганизмов,изучающая отношения между их группами.
Классификация-распределение множества организмов по группам.Таксономия-наименование групп организмов и установлении границ в отношении подчинения.цель классификации:установить степень сходства и революционную взаимосвязь.Царство Протисты-название с 1866года Э. Геккель.Monera-доклеточные формы,Protista-одноклеточные про и эукариоты,Fungi-грибы,Plantae-растения,Animalia-животные.Вид-основная единица систематики.группа близких между собой организмов,имеющие общий корень происхождения ,характеризующийся определенными морфологическими,биохимическими,физиологическими и генетическими признаками,обособленные отбором от других видов и живущие в определенной среде обитания.Классификация делится на :искусственную(ключевая) и естественная (филогенетическая).Цель естественной -объединение родственных форм ,связанных с общностью происхождения,установляемые иерархического подчинения от групп..часто мы пользуемся искусственной класификацией.она объединяет организмы на основе сходства морфологических,биохимических,цитологических признаков.И благодаря этому они объединяются в таксономические группы.Д. Берджи написал главный определитель бактерий по принципу искусственной классификации.Признаки:Морфология,агрегаты,жгутики,споры,окраска по граму,отношение к кислороду,как получают энергию,рост в пределах ph,пит вещества,место обитания,симбиотическая или паразитическая формы,клеточные включения,юкомпоненты клеточной стенки,серологическая характеристика,состав днк,чувствительность к антибиотикам.
Штамм-происхождение из конкретного источника.Варианты-чисто микробное понятие,когда учитывается нестабильность признаков.
7 вопрос.отличие про и эукариот:.Ядро:у прокариот "прокариотное ядро" ,нуклеоид,нет мембраны,.нуклеиновые кислоты:отдельные молекулы,нет гистонов.состав мембран:внутренняя мембрана у цианобактерий,мембраны имеют инвагинации.нет стеролов.Дыхательная система:находится в мезосомах или в мембране.Рибосомы:70 S.Движения цитоплазмы не обнаружено.Состав клеточной стенки :пептидогликан,муреин,а археи в клеточной стенки содержат много белков.1 или несколько фибрилл(жгутики).Вакуоли встречаются очень редко,но есть газовые вакуоли в клетках синезеленых бактерий.положительное действие антибиотиков.предельная температура около 90 градусов у вегетативных клеток,100 градусов у термофилов,выше могут выдержаь только споры.есть бактериохлорофилл,осуществляют фотосинтез,половой процесс представлен фрагментарным процессом.1 кольцевая гаплоидная хромосома.цитоплазматическая днк кольцевая(плазмида) ,не окруженная мембраной.Эукариоты:есть ядро и ядерная мембрана,днк организована в хромосомы,есть гистоны.в митохондриях расположена дых система,80 S рибосомы.есть движение цитоплазмы.клеточная стенка есть у растений и содержит целлюлозу,хитин.жгутиков 11,вакуоли есть у растений у животных нет ,но могут вакуолизироваться в случае прионной болезни.действие антибиотиков отрицательно.предел температуры около 60 градусов.у растений есть фотосинтез.половой процесс в виде мейоза,хромосомы диплоидные больше одной линейные.Запасные вещества прокариот представлены полисахаридами , липидами , полипептидами , полифосфатами , отложениями серы Из полисахаридов в клетках откладываются гликоген , крахмал и крахмалоподобное вещество - гранулеза . Последняя - специфический запасной полисахарид анаэробных споровых бактерий группы клостридиев . Названные полисахариды построены из остатков глюкозы . В неблагоприятных условиях они используются в качестве источника углерода и энергии.
8 вопрос.Изменение условий внешней среды оказывает воздействие на жизнедеятельность микроорганизмов. Физические, химические, биологические факторы среды могут ускорять или подавлять развитие микробов, могут изменять их свойства или даже вызывать гибель.
К факторам среды, оказывающим наиболее заметное действие на микроорганизмы, относятся влажность, температура, кислотность и химический состав среды, действие света и других физических факторов.
Влажность
Микроорганизмы могут жить и развиваться только в среде с определенным содержанием влаги. Вода необходима для всех процессов обмена веществ микроорганизмов, для нормального осмотического давления в микробной клетке, для сохранения ее жизнеспособности. У различных микроорганизмов потребность в воде не одинакова. Бактерии относятся в основном к влаголюбивым, при влажности среды ниже 20 % их рост прекращается. Для плесеней нижний предел влажности среды составляет 15%, а при значительной влажности воздуха и ниже. Оседание водяных паров из воздуха на поверхность продукта способствует размножению микроорганизмов.
При снижении содержания воды в среде рост микроорганизмов замедляется и может совсем прекращаться. Поэтому сухие продукты могут храниться значительно дольше продуктов с высокой влажностью. Сушка продуктов позволяет сохранять продукты при комнатной температуре без охлаждения.
Некоторые микробы очень устойчивы к высушиванию, некоторые бактерии и дрожжи в высушенном состоянии могут сохраняться до месяца и более. Споры бактерий и плесневых грибов сохраняют жизнеспособность при отсутствии влаги десятки, а иногда и сотни лет.
Температура
Температура — важнейший фактор для развития микроорганизмов. Для каждого из микроорганизмов существует минимум, оптимум и максимум температурного режима для роста. По этому свойству микробы подразделяются на три группы:
психрофилы - микроорганизмы, хорошо растущие при низких температурах с минимумом при -10-0 °С, оптимумом при 10-15 °С;
мезофилы - микроорганизмы, для которых оптимум роста наблюдается при 25-35 °С, минимум — при 5-10 °С, максимум — при 50-60 °С;
термофилы - микроорганизмы, хорошо растущие при относительно высоких температурах с оптимумом роста при 50-65 °С, максимумом — при температуре более 70 °С.
Большинство микроорганизмов относится к мезофилам, для развития которых оптимальной является температура 25-35 °С. Поэтому хранение пищевых продуктов при такой температуре приводит к быстрому размножению в них микроорганизмов и порче продуктов. Некоторые микробы при значительном накоплении в продуктах способны привести к пищевым отравлениям человека. Патогенные микроорганизмы, т.е. вызывающие инфекционные заболевания человека, также относятся к мезофилам.
Низкие температуры замедляют рост микроорганизмов, но не убивают их. В охлажденных пищевых продуктах рост микроорганизмов замедленно, но продолжается. При температуре ниже О °С большинство микробов прекращают размножаться, т.е. при замораживании продуктов рост микробов останавливается, некоторые из них постепенно отмирают. Установлено, что при температуре ниже О °С большинство микроорганизмов впадают в состояние, похожее на анабиоз, сохраняют свою жизнеспособность и при повышении температуры продолжают свое развитие. Это свойство микроорганизмов следует учитывать при хранении и дальнейшей кулинарной обработке пищевых продуктов. Например, в замороженном мясе могут длительно сохраняться сальмонеллы, а после размораживания мяса они в благоприятных условиях быстро накапливаются до опасного для человека количества.
При воздействии высокой температуры, превышающей максимум выносливости микроорганизмов, происходит их отмирание. Бактерии, не обладающие способностью образовывать споры, погибают при нагревании во влажной среде до 60-70 °С через 15-30 мин, до 80-100 °С — через несколько секунд или минут. У спор бактерий термоустойчивость значительно выше. Они способны выдерживать 100 °С в течение 1-6 ч, при температуре 120-130 °С споры бактерий во влажной среде погибают через 20-30 мин. Споры плесеней менее термостойки.
Тепловая кулинарная обработка пищевых продуктов в общественном питании, пастеризация и стерилизация продуктов в пищевой промышленности приводят к частичной или полной (стерилизация) гибели вегетативных клеток микроорганизмов.
При пастеризации пищевой продукт подвергается минимальному температурному воздействию. В зависимости от температурного режима различают низкую и высокую пастеризацию.
Низкая пастеризация проводится при температуре, не превышающей 65-80 °С, не менее 20 мин для большей гарантии безопасности продукта.
Высокая пастеризация представляет собой кратковременное (не более 1 мин) воздействие на пастеризуемый продукт температуры выше 90 °С, которая приводит к гибели патогенной неспороносной микрофлоры и в то же время не влечет за собой существенных изменений природных свойств пастеризуемых продуктов. Пастеризованные продукты не могут храниться без холода.
Стерилизация предусматривает освобождение продукта от всех форм микроорганизмов, в том числе и спор. Стерилизация баночных консервов проводится в специальных устройствах — автоклавах (под давлением пара) при температуре 110-125°С в течение 20-60 мин. Стерилизация обеспечивает возможность длительного хранения консервов. Молоко стерилизуется метолом ультравысокотемпературной обработки (при температуре выше 130 °С) в течение нескольких секунд, что позволяет сохранить все полезные свойства молока. Реакции среды:
Жизнедеятельность микроорганизмов зависит от концентрации водородных (Н+) или гидроксильных (ОН-) ионов в субстрате, на котором они развиваются. Для большинства бактерий наиболее благоприятна нейтральная (рН около 7) или слабощелочная среда. Плесневые грибы и дрожжи хорошо растут при слабокислой реакции среды. Высокая кислотность среды (рН ниже 4,0) препятствует развитию бактерий, однако плесени могут продолжать расти и в более кислой среде. Подавление роста гнилостных микроорганизмов при подкислении среды имеет практическое применение. Добавление уксусной кислоты используется при мариновании продуктов, что препятствует процессам гниения и позволяет сохранить продукты. Образующаяся при квашении молочная кислота также подавляет рост гнилостных бактерий. Концентрация соли и сахара:
Поваренная соль и сахар издавна используются для повышения стойкости продуктов к микробной порче и лучшей сохранности пищевых продуктов.
Повышение содержания растворенных веществ (соли или сахара) в питательной среде сказывается на величине осмотического давления внутри микроорганизмов, вызывает их обезвоживание. При повышении концентрации поваренной соли в субстрате более 3-4 % размножение многих, в том числе гнилостных, микроорганизмов замедляется, при концентрации более 7-12% — прекращается.
Некоторые микроорганизмы нуждаются для своего развития в высоких концентрациях соли (20 % и выше). Их называют солелюбивыми, или галофилами. Они могут вызывать порчу соленых продуктов.
Высокие концентрации сахара (выше 55-65 %) прекращают размножение большинства микроорганизмов, это используется при приготовлении из плодов и ягод варенья, джема или повидла. Однако эти продукты тоже могут подвергаться порче в результате размножения осмофильных плесеней или дрожжей. Свет:
Некоторым микроорганизмам свет необходим для нормального развития, но для большинства из них он губителен. Ультрафиолетовые лучи солнца обладают бактерицидным действием, т. е. при определенных дозах облучения приводят к гибели микроорганизмов. Бактерицидные свойства ультрафиолетовых лучей ртутно-кварцевых ламп используют для дезинфекции воздуха, воды, некоторых пищевых продуктов. Инфракрасные лучи тоже могут вызвать гибель микробов за счет теплового воздействия. Воздействие этих лучей применяют при тепловой обработке продуктов. Негативное воздействие на микроорганизмы могут оказывать электромагнитные поля, ионизирующие излучения и другие физические факторы среды. Химические факторы:
Некоторые химические вещества способны оказывать на микроорганизмы губительное действие. Химические вещества, обладающие бактерицидным действием, называют антисептиками. К ним относятся дезинфицирующие средства (хлорная известь, гипохлориты и др.), используемые в медицине, на предприятиях пищевой промышленности и общественного питания.
Некоторые антисептики применяются в качестве пищевых добавок (сорбиновая и бензойная кислоты и др.) при изготовлении соков, икры, кремов, салатов и других продуктов. Биологические факторы:
Между различными микроорганизмами могут устанавливаться разные взаимоотношения: симбиоз- взаимовыгодные отношения; метабиоз — жизнедеятельность одного за счет другого без принесения вреда; паразитизм — жизнедеятельность одного за счет другого с причинением ему вреда; антагонизм — один из видов микроорганизмов угнетает развитие другого, что может привести к гибели микробов. Например, развитие молочнокислых бактерий угнетает рост гнилостных, эти антагонистические взаимоотношения используют при квашении овощей или для поддержания нормальной микрофлоры в кишечнике человека.
Антагонистические свойства некоторых микроорганизмов объясняются способностью их выделять в окружающую среду вещества, обладающие антимикробным (бактериостатическим, бактерицидным или фунгицидным) действием, - антибиотики. Антибиотики продуцируются в основном грибами, реже бактериями, они оказывают свое специфическое действие на определенные виды бактерий или грибов (фунгицидное действие). Антибиотики применяются в медицине (пенициллин, левомицетин, стрептомицин и др.), в животноводстве в качестве кормовой добавки, в пищевой промышленности для консервирования пищевых продуктов (низин).
Антибиотическими свойствами обладают фитонциды — вещества, обнаруженные во многих растениях и пищевых продуктах (лук, чеснок, редька, хрен, пряности и др.). К фитонцидам относятся эфирные масла, антоцианы и другие вещества. Они способны вызывать гибель патогенных микроорганизмов и гнилостных бактерий.
9 вопрос.Вода. Основная составная часть бактериальной клетки, приходящаяся нга воду, — 75—85 %, сухое вещество составляет 15—25 %. Часть воды находится в свободном состоянии, а часть — в связанном. Связанная вода является структурным растворителем. Свободная вода служит дисперсионной средой для коллоидов, растворителем для кристаллических веществ, источником водородных и гидроксиль-ных ионов. (Например, гидролитические процессы расщепле-
Ведущая роль принадлежит четырем органогенам — кислороду, водороду, углероду и азоту. В процентном отношении к сухому веществу бактерии содержат: углерода — 45—55, азота — 8—15, кислорода — 30, водорода-—6—8. Соответственно дрожжи содержат: углерода — 49, азота— 12, кислорода—-.31, водорода — 6%. В микроскопических грибах, углерода — 47, азота — 5, кислорода — 40, водорода — 6%. Минеральные вещества. Кроме органогенов в микробных клетках находятся так называемые зольные элементы — минеральные вещества, составляющие от 3 до 10 % сухого вещества микроорганизмов.
Белки — это высокомолекулярные азотсодержащие органические соединения, молекулы которых построены из аминокислотных остатков, соединенных между собой ковалент-ными пептидными связями.
Нуклеиновые кислоты представляют собой высокомолекулярные биологические полимеры, построенные из мононук-леотидов. Особенно характерно для них содержание фосфора (8—10%) и азота (15—16%), они также содержат углерод, кислород и водород.
Углеводы. В бактериях их содержится 12—18% от сухого вещества. Это многоатомные спирты (сорбит, маннит, дуль-цит); полисахариды (гексозы, пентозы, гликоген, декстрин); моносахариды (глюкоза, глюкуроновая кислота и др.). Углеводы выполняют энергетическую роль в микробной клетке. Липиды-истинные жиры ,липоиды-жироподобные вещества .Факторы роста-это биологически активные вещества( (ряд аминокислот, витамины, пуриновые и пиримидиновые основания, стерины и др.), в отсутствии к-рых мн. микроорганизмы не растут даже на питат.средах, содержащих необходимые источники энергии, углерода и азота. М. ф. р. оказывают действие в ничтожных количествах. Неспороносные бактерии (Pseudomonas, Mycobacterium), мн. плесневье грибы (Aspergillus, Penicillium) и др. микроорганизмы не нуждаются в М. ф. р., так как способны их синтезировать Аминокислоты необходимы для биосинтеза белка, пурнновые и пиримидиновые основания - для образования нуклеиновых к-т. Нек-рые виды дрожжей, молочнокислые бактерии не растут на питат. средах без биотина, тиамина пантотеновой и никотиновой к-т, пиридоксипа и др. Считали, что нек-рые болезнетворные микроорганизмы растут только на средах содержащих кровь или eё сыворотку, асцитическую жидкость, молочную сыворотку, дрожжевой автолизат. Оказалось, что эти микробы могут расти и без таких веществ, если к питат. среде добавить соответствующие М. ф. р. Если М. ф. р. имеют сложное химич. строение, то потребность в них у разных микроорганизмов может варьировать. Т. н. дикие формы микроорганизмов, способные к синтезу М. ф. р., паз. прототрофными. Воздействуя на них мутагенами, можно получить мутанты, нуждающиеся в том или ином М. ф. р. Их называют ауксотрофными или дефицитными мутантами и применяют для количеств, определения витаминов, аминокислот и т. д., а также при селекции мутантов, образующих повышенные кол-ва этих веществ. Способность к синтезу М. ф. р. может определять характер взаимоотношений между организмами. Так, если определённый вид дрожжей не растёт из-за отсутствия в питат. среде М. ф. р., то подсев и размножение вида, синтезирующего их, приведёт к одновременному росту обоих видов. У нек-рых насекомых и ракообразных имеются микроорганизмы-симбионты, размножающиеся в кишечнике или особых органах и снабжающие организм хозяина различными витаминами, аминокислотами и т. п. Микроорганизмы, обитающие в рубце и кишечнике жвачных, а также в кишечнике др. животных и человека, выполняют ту же функцию. Основные принципы культивирования микрорганизмов на питательных средах:1.использование всех необходимых для микробов питательных веществ.2.оптимальные температуры,pH,концентрация ионов, степень насыщения кислородом, газовый состав и давление. Микроорганизмы культивируют на питательных средах при оптимальной температуре и в термостатах,обеспечивающие условия инкубации. По температурному оптимуму выделяют:Психрофилы-растут при температуре ниже 20 градусов.2.Мезофилы-растут в диапазоне от 20-45 градусов.3.Термофилы растут при температуре выше плюс 40 градусов. Среды выделяют жидкие,полужидкие(0,3-0,7 % агара),плотные(1,5-3 % агара).
10 вопрос: Подобно прочим формам жизни, бактерии нуждаются в одних и тех же макроэлементах — С, Н, О, N, P, S, К, Са, Mg, Fe. Микроэлементы (следовые элементы) — Mn, Mo, Zn, Си, Со, Ni, Va, В, С], Na, Se, Si, Wo — не нужны каждому организму, но бактериям они необходимы для синтеза коферментов либо поддержания специфического тина метаболизма. Например, для оптимального роста некоторые бактерии нуждаются в высоких концентрациях Na+; их называют галофилами [от греч. hals, соль]. Помимо источников углерода, энергии и элементов минерального питания, многие микроорганизмы нуждаются в некоторых дополнительных веществах, называемых факторами роста. Количественная потребность в питательных элементах и их содержание у различных бактерий варьируют, но принципиально химический состав бактериальной клетки сходен с другими живыми клетками (исключением является отсутствие у бактерий стеролов).
Ферменты: 1) снижают энергию активации хим.реакций, 2) увеличивают скорость реакций, 3) обуславливают узнавание субстрата (специфичность) и стерические свойства (ключ-замок). Каждый фермент может катализировать реакцию в обоих направлениях.
Классификация ферментов: 1) оксидоредуктазы – перенос электронов, 2) трансферазы – перенос групп, 3) гидролазы – перенос групп на молекулу Н2О, 4) лиазы - образования и разрыва двойных связей С, 5) изомеразы – перенос грцпп внутри молекулы, 6) лигазы – сшивают С-С, C-S, C-O,C-N; распад АТФ.
Пассивный транспорт - протекает без затраты энергии, вещества перемещаются от более высокой концентрации к более низкой.
Простая диффузия. Проникновение веществ носит неспецифический характер и целиком зависит от размеров молекул и их липофильности. Скорость подобного переноса незначительна.
Облегчённая диффузия. Механизм транспорта носит аналогичный характер, но проникновение облегчают помощники — специфические мембранные белки-пермеазы, способствующие прохождению различных молекул через ЦПМ. Транспорт сопровождается образованием комплекса «вещество-пермеаза». После преодоления ЦПМ комплекс диссоциирует, а перме-аза используется для последующего «проведения» других молекул. Подобный тип транспорта реализуется по градиенту концентрации и характерен для эукариотов при поглощении сахаров.
Активный транспорт — перенос вещества через клеточную мембрану, протекающий против градиента концентрации из области низкой концентрации в область высокой, т. е. с затратой свободной энергии организма. В большинстве случаев, но не всегда, источником энергии служит энергия макроэргических связей АТФ.
Механизм активного транспорта лучше всего изучен для натрий-калиевого насоса (Na+/K+-нaсоса) — транспортного процесса, который выкачивает ионы натрия через мембрану клетки наружу и в то же время закачивает в клетку ионы калия. Этот насос отвечает за поддержание различной концентрации ионов натрия и калия по обе стороны мембраны, а также за наличие отрицательного электрического потенциала внутри клеток.
Мембранный транспорт веществ различается также по направлению их перемещения и количеству переносимых данным переносчиком веществ:1) Унипорт — транспорт одного вещества в одном направлении в зависимости от градиента 2) Симпорт — транспорт двух веществ в одном направлении через один переносчик. 3) Антипорт — перемещение двух веществ в разных направлениях через один переносчик.
Транспорт, обусловленный фосфорилированием — энергозависимый процесс, используемый при утилизации углеводов. Основной механизм транспорта связан с фосфорилированием субстрата, что делает невозможным его выход из клетки. Первоначально происходит фосфорилирование мембранного фермента (так называемый фермент 2, или Ф2) в цитоплазме за счёт фосфоенолпирувата. Фосфорилированный фермент связывает углевод (например, глюкозу или маннозу) на поверхности ЦПМ и транспортирует его в цитоплазму. Затем комплекс диссоциирует с высвобождением углевода, связанного с фосфатной группой. За счёт фосфорилирования углевод аккумулируется в клетке и не способен выходить из неё.
11 вопрос. Пищевые потребности микроорганизмов. Способы питания микроорганизмов. Источники углерода. Источники энергии. Доноры электронов.
Пищей принято называть любое вещество, которое попав в организм служит источником Е и/или пластическим материалом для синтеза молекул, используемых для роста.
По типу питания существует несколько типов классификации:
1) Голозои(тв.пища) / голофиты (жидк.растворен.пища). Мко относятся к голофитам(питаются растворенной пищей). Мко секретируют ферменты (литические), они действуют на целлюлозное волокно(полимер), образуется глюкоза – она может поглощаться мко. Род целлюлозоразлагающих бактерий – Cytophaga. При ферментативном гидролизе целлюлозы бактериями, обитающими в ЖКТ жвачных животных образуется целлобиоза. Она расщепляется бактериальным ферментом β-глюкозидазой (целлобиазой) до глюкозы, что обеспечивает усвоение жвачными целлюлозной части биомассы.
Протеазы – ферменты, расщепляющие белки. Липазы – ферменты, расщепляющие жиры.
2) живое орг.в-во / мертвое орг.в-во
Хищники (исп-ют жив.орг.в-во,быстро) – Паразиты (исп-ют жив.орг.медленно) – Сапрофиты (исп-ют мертв.орг.). Мко относятся ко всем трем. Род Bdellvibrio(подвижный вибрион) питается грам- бактериями (род Pseudomonas – очищают воду)
орг.С (живой) н-р белки |
орг. С (мертв) |
орг. С + СО2 (неорг) |
СО2 (неорг) |
Паратрофы облигатные→факульт-е → паразиты паразиты -риккетсии -микоплазмы -хламидии -Bdellvibrio -вирусы -прионы
|
Метатрофы спрофиты → мезотрофы/олиготрофы -псевдомон -псевдомо -клостридии |
Миксотрофы Цианобактерии →
|
Автотрофы на свету / в темноте |
3) источник С
Вода. 80% веса бактерий. Если сдвинуть рн клетки от 7,2 до 7,4, то можно имитировать деление клетки(В.П.Скулачев) – среда для реакций, ф-ция регуляторная.
Основные соед-я, которые усваивает бактериальная клетка: углеводы, белки, амк, жирные к-ты, минер.соли, витамины и др. Некоторые мко могут разложить(утилизировать) мыло, парафин, карбоновые к-ты.
Макроэл-ты бактер.клетки: С, Н, О, N, P, S, K, Ca, Fe. Микроэл-ты бак.клетки: Мо, Zn, Cu, Co, Ni, B, Cl, Na, Se, Si, Wo…-необязательны всегда явл-ся частью системы поддерж-й микробный гомеостаз. Либо для коферментов, либо для специфического метаболизма. Дополнит.в-ва – необходимы некотор.мко, которые сами не способны их синтезировать (Ауксотрофы–бактерия не может синтезировать что-то): гистидин, пурин. Мед.значимые формы.
В зависимости от источника углерода микроорганизмы делятся на: автотрофы (сами себя питающие), которые используют углерод из неорганических соединений (углекислого газа и карбонатов); гетеротрофы (питаются за счет других) – используют углерод из органических соединений.
В зависимости от источника энергии различают: фототрофы – микроорганизмы, которые в качестве источника энергии используют энергию солнечного света; хемотрофы – энергетическим материалом для этих микроорганизмов являются разнообразные органические и неорганические вещества.
В зависимости от источника электронов (природы окисляемого субстрата) микроорганизмы делятся на: литотрофы – окисляют неорганические вещества и за счет этого получают энергию; органотрофы – получают энергию путем окисления органических веществ.
Восстановленные соединения являются донорами е- и [Н+]. H2S(отдает е-)
Кроме углерода, важнейшим элементом питательной среды является азот. Автотрофы обычно используют азот из минеральных соединений, а гетеротрофы, кроме неорганических соединений азота, используют аммонийные соли органических кислот, аминокислоты, пептоны и другие соединения. Некоторые гетеротрофы усваивают атмосферный азот (азотфиксаторы).
Источник Е |
Донор е- |
Источник С |
Способ существ-я |
Примеры |
Окислит-восстано вит р-ции (Хемо-) |
Неорг. соед-я (лито-) |
СО2 (авто-) |
ХЛА |
Нитрифицирующие, тионовые |
орг (гетеро-) |
ХЛГ |
Водородные, метаногены |
||
Органич. соед-я (органо-) |
СО2 (авто-) |
ХОА |
Метилотрофы |
|
орг (гетеро-) |
ХОГ |
Больш-во прокар +грибы, жив-е |
||
Свет (Фото-) |
Неорг. соед-я (лито-) |
СО2 (авто-) |
ФЛА |
Цианобак, пурпур, зелен +раст-я |
орг (гетеро-) |
ФЛГ |
Некот цианоб и пурпур,зеленые |
||
Органич. соед-я (органо-) |
СО2 (авто-) |
ФОА |
Некоторые пурпурные бактерии |
|
орг (гетеро-) |
ФОГ |
Некот.циано,пурп, галобактерии |