

3. Развитие и созревание мозга Дж. Шаде, д. Форд [Морфогенез и гистогенез]*
* Шаде Дж., Форд Д. Основы неврологии. М.:Мир, 1976. С. 14-38.
(Тематические заголовки в квадратных скобках даны редактором-составителем.)
Морфогенез
Рис. 3.1. Мозг человека и остальных позвоночных развивается из эмбриональной эктодермы. При развитии человеческого эмбриона нервная система, формирующаяся из эктодермы, представлена двумя структурами: первичной полоской (7) и нотохордом (2). В конце третьей недели эмбрион представляет собой овальный диск около 1,5 см в длину. Длинная ось диска определяется наличием в его заднем квадранте первичной полоски, которую можно рассматривать как область продольного утолщения. Непосредственно кпереди (рострально) от первичной полоски находится другое утолщение эктодермы, называемое гензеновским, или первичным, узелком. Центр этого узелка втягивается благодаря движению поверхностных клеток и образует углубление, бластопор (3). Затем из гензеновского узелка в ростральном направлении между эктодермой и энтодермой мигрирует тяж клеток, который образует нотохорд (2). В переднем отделе эмбрионального диска этот тяж прикрепляется к прохордальной пластинке (4). В процессе развития происходит дальнейшее втягивание бластопора, переходящего в нотохорд и образующего продолжение амниотической полости, называемое нотохордальным каналом.

Рис. 3.1. Модель дорсальной части 18-дневного эмбриона, на которой видна медуллярная пластинка. Типичные срезы, сделанные по линиям А, Б, В, представлены соответствующими схемами:
5 – эмбриональная эктодерма; 6 – нервная пластинка; 8 – желточный мешок; 9 – эктодерма амниона; 10 – эмбриональная мезодерма; 11 – энтодерма. Остальные объяснения см. в тексте
Рис. 3.2. Эктодерма (5), покрывающая нотохорд, затем утолщается, что вызывается, как полагают, влиянием нотохорда и прохордальной мезодермы, к которой прикрепляется нотохорд. Утолщение эктодермы формирует нервную пластинку (6). В дальнейшем нервная пластинка прогибается с образованием сначала нервной бороздки (7), а в конечном итоге – длинной полой трубки, лежащей непосредственно под поверхностью эктодермы, от которой она отделяется. Еще в 1895 г. было высказано предположение (Фуа), что превращение нервной пластинки в полую трубку вызывают силы, присущие самой пластинке. Образование валиков происходит даже в том случае, если пластинку извлекают из эмбриона. Сливаясь, валики образуют нервную трубку (8). Свободные эпидермальные слои, от которых отделилась нервная трубка, также соединяются и, сливаясь, образуют нейроэктодермальные соединения над верхним краем вновь образованной нервной трубки. Сформированная нервная трубка расширяется на переднем конце, на месте формирования будущего головного мозга. Более тонкая каудальная часть нервной трубки преобразуется в спинной мозг.

Рис. 3.2. Различные стадии образования нервной трубки из нервной пластинки и сопутствующее формирование ее моторного, сенсорного и вегетативного отделов:
1 – амнион; 2 – мезодерма; 3 – сомит; За – дерматом; 36 – миотом; 12 – мантийный слой; 13 – краевой слой; 14 – спинальный ганглий; 15 – передний корешок спинного мозга; 16 – клетки переднего рога спинного мозга; 17 – симпатический ганглий; 18 – преганглионарная ветвь; 19 – постганглионариая ветвь; 20 – висцеральная ветвь; 21 – крыловидная (сенсорная) пластинка; 21а – крыша; 22 – пограничная бороздка (sulcus limitans); 23 – базалъная (моторная) пластинка; 2З а – дно; 24 – мышца. Остальные объяснения см. в тексте
Первой замыкается та часть нервной трубки, которая образует задний мозг (rhombencephalon). При замыкании бороздки в каудальном направлении формируется та часть нервной трубки, которая соединяется со спинным мозгом. Замыкание нервной бороздки в ростральном направлении происходит более или менее одновременно с формированием сегментов спинного мозга. Оно сначала ведет к образованию среднего мозга (mesencеphalon), а затем переднего мозга (prosеncephalon) – надсегментарных образований, лежащих ростральнее заднего мозга. Последним замыкается небольшое отверстие на мозговом конце, называемое передним нейропором <...>.
Показано, что щелевидная форма полости нормальной нервной трубки зависит от наличия подлежащего нотохорда (рис. 3.2, А, 4). По мере того как нервная трубка (6) замыкается сверху, эктодермальные клетки боковых краев нервной бороздки выталкиваются в сторону, образуя продолговатый тяж клеток по обеим сторонам трубки, нервный гребень (10). Процесс формирования нервной трубки называется нейруляцией. По окончании нейруляции эмбрион, лишенный ранее оси симметрии, приобретает билатеральную симметрию. Теперь уже отчетливо выражена цефализация и различимы головной и хвостовой концы эмбриона.
Какие силы вызывают образование нотохорда, нервной трубки и их производных? Хотя окончательного ответа на этот вопрос еще не существует, полагают, что всем морфологическим изменениям эмбриона предшествует установление метаболического градиента, который характеризуется определенной цефалокаудальной ориентацией. Полагают также, что эти метаболические различия связаны с существованием химических градиентов. Так, у эмбрионов рыб и амфибий отмечается высокая концентрация гликогена в той презумптивной эктодерме, из которой формируется нервная трубка, тогда как уровень гликогена в нотохорде и энтодерме очень низок. Помимо этого, существует цефалокаудальный градиент содержания гликогена. Установлено, что в тех областях, где протекают процессы морфогенеза, синтез белка отличается особой интенсивностью. Столь высокая интенсивность синтеза белка сочетается с увеличением концентрации нуклеопротеидов, количество которых уменьшается по мере того, как дифференцировка в данной области подходит к концу. Затем концентрация нуклеопротеидов может возрастать в какой-либо другой области, снова, по-видимому, несколько предшествуя морфогенезу. Нервная пластинка – это одна из областей, которая характеризуется высокой концентрацией рибонуклеопротеидов с наличием цефалокаудального градиента, что предположительно указывает на возрастание синтеза белка в этой области, как это, возможно, и происходит при замыкании нервной трубки. Упомянутые выше градиенты сочетаются также с градиентами активности дыхательных ферментов. Хотя окончательных подтверждений взаимозависимости упомянутых выше факторов еще не получено, кажется вполне логичной концепция связи между цефалокаудальными градиентами и образованием РНК и белка в ходе развития нервной трубки.
Как только сформировалась нервная трубка (рис. 3.2, В, 8), клетки нервного гребня (10) мигрируют кнаружи и дают начало спинальным ганглиям (рис. 3.2, Д, 14), периферическим постганглионарным нейронам висцеромоторной (симпатической) системы, шванновским клеткам, которые формируют липидные (миелиновые) оболочки периферических нервов, поддерживающим (сателлитным) клеткам нейронов спинальных ганглиев, а помимо этого, клеткам, образующим два внутренних листка оболочек, окутывающих мозг. Происходят изменения и в самой нервной трубке благодаря пролиферации клеток, выстилающих ее центральный канал. Пролиферативной зоной здесь служит слой эпендимы, который дает начало нейробластам, превращающимся в нервные клетки, и спонгиобластам, которые становятся глиальными клетками – опорными элементами нервной системы.
В результате различной скорости деления и накопления клеток центральный канал суживается двумя продольно ориентированными клеточными пластинками (рис. 3.7). Расположенные более дорсально крыловидные пластинки (рис. 3.2, Е,21) функционально могут рассматриваться как сенсорные в спинном мозге и нижних отделах мозгового ствола, тогда как вентральные, или базальные, пластинки (23) выполняют двигательные функции. Продольная бороздка на границе пластинок называется sulcus limitans (22). Обе крыловидные пластинки соединяются сверху посредством крыши (21 а), а базальные пластинки – внизу с помощью дна (23а). В отделах головного мозга, лежащих ростральнее продолговатого мозга, из крыловидной пластинки формируется множество структур, которые не являются сенсорными, а выполняют моторные, ассоциативные или релейные функции (например, полушария мозга, промежуточный мозг, мозжечок или ядра моста). Надо подчеркнуть, что все двигательные системы низших уровней (нейроны передних рогов спинного мозга, двигательные ядра черепномозговых нервов), иннервирующие мышцы, железы и т. д. происходят из базальной пластинки. Поскольку ростральнее среднего мозга не существует ни крыловидной, ни базальной пластинок, то нельзя и соотносить моторные или сенсорные компоненты функций с этими структурами.
Рис. 3.3. Длинная полая нервная трубка, сформировавшись, быстро подвергается дальнейшим изменениям. На ранней стадии развития (рис. 3.3, А) она подразделяется на длинную каудальную трубку, которая образует спинной мозг (4) и более широкие, короткие ростральные сегменты, которые превращаются, в головной мозг. Головной конец нервной трубки вскоре подразделяется на три расширения, первичные мозговые пузыри (А). Полости этих пузырей сохраняются в мозге взрослого в видоизмененной форме и образуют полости желудочков и сильвиева водопровода.

Рис. 3.3. Развивающийся мозг:
А – формирование первичных пузырей (до 4-й недели эмбрионального развития); Б – Е – формирование вторичных пузырей; Б, В – конец 4-й недели; Г – 6-я неделя; Д – 8–9-я недели, завершающиеся формированием основных отделов мозга (Е) к 14-й неделе; За – перешеек ромбовидного мозга; 7 – конечная пластинка. Остальные объяснения см. в тексте
Самым ростральным отделом нервной трубки является prosencephalon, или передний мозг (1), за ним следует mesencephalon, или средний мозг (2); третий, наиболее каудально расположенный сегмент образует rhombencephalon, или задний мозг(3). В последующем развитии prosencephalon делится на telencephalon, или конечный мозг(полушария большого мозга к некоторые базальные ядра, 5), и diencephalon, или промежуточный мозг(6). С каждой стороны промежуточного мозга вырастает глазной пузырь, формирующий нервные элементы глаза. Глазной бокал, образованный этим выростом, вызывает изменения в лежащей непосредственно над ним эктодерме, что приводит к возникновению хрусталика (ср. рис. 3.5). Средний мозг сохраняется как единое целое, но в процессе развития в нем происходят значительные изменения, связанные с образованием специализированных рефлекторных центров, имеющих отношение к зрению, слуху, а также к тактильной, болевой и температурной чувствительности.
Ромбовидный мозг подразделяется на metencephalon, или задний мозг, включающий мозжечок(8) и мост(9), и myelencephalon, или продолговатый мозг (medulla oblongata, 10).
Изгибы нервной трубки
Описанные выше изменения нервной трубки сопровождаются образованием нескольких изгибов, что обусловлено разной скоростью роста отдельных ее частей. Конечным итогом этих изменений будет загиб теленцефало-диенцефального конца вперед в области среднемозгового изгиба и под прямым углом по отношению ко всей остальной нервной трубке (ср. рис. 3.5). Поскольку номенклатура отделов мозга основана преимущественно на его описаниях у животных, у которых отсутствует подобный изгиб между передним и средним мозгом, возникли некоторые затруднения в терминологии. Однако они могут быть частично преодолены, если мысленно представить себе человека в позе четвероногого животного (рис. 3.4) и, исходя из этого, использовать различные термины для обозначения отделов мозга.

Рис. 3.4. Наиболее принятые анатомические обозначения:
А – рисунок, изображающий человека в положении, соответствующем положению тела четвероногого, так что мозг и корешки спинного мозга располагаются таким образом, что передние и задние, ростральные и каудальные отделы этих структур могут быть сопоставлены с их расположением у животных: Б, В – общепринятые плоскости сечения мозга при анатомических и патоморфологических исследованиях; а – срединная (сагиттальная) плоскость; 6 – парасагиттальная и в – фронтальная (коронарная) плоскости; г – плоскость, лежащая под углом 15–20° к горизонтальной плоскости
Рис. 3.5 и 3.6. По мере развития нервной трубки различные изгибы формируются отчасти в результате более быстрого роста некоторых ее отделов. При этом изгибы возникают из-за того, что то место, где передний мозг прикреплен к вершине букофарингеальной мембраны, представляет собой сравнительно неподвижную точку по отношению к остальной нервной трубке, меняющей свое положение.

Рис. 3.5. Развивающийся мозг (с 3-й по 7-ю неделю развития):
Вид сбоку, иллюстрирующий изгибы в среднемозговой (а), шейной (б) областях мозга, а также в области моста (в): 1 – глазной пузырь, глаз; 2 – передний мозг; 3 – средний мозг; 4 – задний мозг; 5 – слуховой пузырек; 6 – спинной мозг; 7 – промежуточный мозг; 8 – конечный мозг; 9 – ромбическая губа. Римскими цифрами обозначены места отхождения черепномозговых нервов
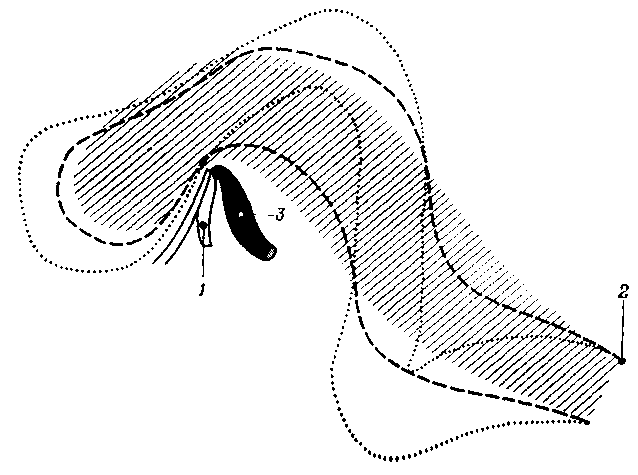
Рис. 3.6. Схема, иллюстрирующая изменения положения нервной трубки в процессе формирования ее изгибов:
Изгибы образуются в результате быстрого роста нервной трубки, фиксированной в двух точках – в области букофарингеалъной мембраны (1) и в области шейных сомитов (2); 3 – нотохорд
Таким образом, начиная с 3-й недели, быстрый рост нервной трубки кпереди и вверх вызывает ее изгиб вперед, так что к 4-й неделе уже существуют изгибы в области, соответствующей будущему среднему мозгу (теменной изгиб, а) и, каудальнее, в месте соединения продолговатого и спинного мозга (затылочный изгиб, б). Несколько позже, в течение 5-й недели, начинает формироваться третий изгиб в области моста (мостовой изгиб, в), который имеет направление, противоположное двум предыдущим, что сопровождается еще большим увеличением кривизны среднемозгового изгиба (а). В конце концов все изгибы, за исключением среднемозгового, исчезнут, и сохранится лишь изгиб мозгового ствола почти под прямым углом в области соединения среднего и промежуточного мозга.
Развитие отдельных областей ЦНС
Спинной мозг
Рис. 3.7. На первых стадиях развития нервная трубка на всем протяжении состоит из трех слоев: эпендимального (1), мантийного (2) и краевого (3). Нейробласты мигрируют кнаружи из эпендимального, или герминативного (зачаткового), слоя в мантийный. Некоторые из этих нейробластов служат предшественниками нейронов, другие же, не будучи истинными нейробластами, представляют собой спонгиобласты, которые в дальнейшем дают начало глиальным элементам. В той же части нервной трубки, которая в дальнейшем образует спинной, продолговатый, задний и средний мозг, в мантийном слое (2) формируются четыре параллельные колонки нейронов: две – соответствующие крыловидным пластинкам (4) – расположены дорсолатерально и две, лежащие вентролатерально, соответствуют базальным пластинкам (5). Эти колонки соединены с дорсальной стороны крышей (6), а с вентральной – дном (7); продольная пограничная бороздка (sulcus limitans) (8) отделяет крыловидную пластинку от базальной. Последняя, из которой формируются важнейшие элементы двигательных центров низших отделов мозга, рострально доходит лишь до среднего мозга. Таким образом, конечный и промежуточный мозг целиком развиваются из колонок клеток, относящихся к крыловидной пластинке.

Рис. 3.7. Развитие спинного мозга:
Рост крыловидной и базальной пластинок у эмбрионов на разных стадиях (А – 8мм; Б – 15 мм; В – 35 мм), ведущий к образованию задних (16) и передних (15) рогов серого вещества (12) развитого спинного мозга (Г); 9 – представительство соматической и 10 – висцеральной функциональных областей. Остальные объяснения см. в тексте
В ходе созревания спинного мозга базальная пластинка дает начало клеткам вентральных (передних) рогов (15) и клеткам интермедиолатерального столба (14). По мере того как масса нейробластов в базальной пластинке увеличивается, образуются группы отдельных ядерных скоплений, так что колонка клеток превращается в ряд последовательно расположенных расширений, более толстых в промежутках между сегментами. Такое первичное сегментарное расположение нейробластов заметно уже у эмбрионов на стадии, соответствующей длине 8– 10 мм. На стадиях, соответствующих 8– 15 мм, базальная колонка подвергается вторичной продольной перестройке с образованием трех групп.
I. Вентролатеральное скопление клеток (а), в котором находятся 2–3 дополнительные периферические колонки. Отсюда происходят нейроны, направляющиеся в вентральный (передний) корешок спинного мозга. Те клетки, которые в дальнейшем у взрослых образуют интермедиолатеральную клеточную колонку (а), происходят из дорсальной части этой вентролатеральной группы.
II. Полоски дорсомедиально ориентированных клеток (б), расположенных глубже и медиальнее клеток группы а; аксоны клеток группы б проходят на противоположную сторону в том месте, где будет находиться вентральная (передняя) комиссура спинного мозга.
III. Небольшая группа нейробластов (в), которая, по-видимому, образуется из зачатковых клеток эпендимы несколько позднее и располагается между слоем эпендимы и клетками группы б, эта группа играет важную роль на более ростральных уровнях нервной трубки и, по-видимому, служит источником клеток, формирующих ядра некоторых черепномозговых нервов.
Крыловидная пластинка (4) дает начало сенсорным, релейным и вставочным нейронам. Нейробласты мантийного слоя крыловидной пластинки растут и дифференцируются медленно и к середине второго месяца образуют массивное скопление клеток, лежащее дорсальнее базальной пластинки. Нейробласты, происходящие из той части эпендимы, которая лежит сразу дорсальнее sulcus limitans (8), формируются, по-видимому, быстрее нейробластов более отдаленной дорсальной части крыловидной пластинки, прилежащей к крыше (6). В результате этого клетки в основании крыловидной пластинки в большей степени распространяются в латеральном направлении (рис. 3.7, В). Часть этих клеток продвигается в дальнейшем в дорсальном направлении, огибая более медленно формирующуюся дорсальную группу клеток, которая образуется из крыловидной пластинки (4а).
Уже на 5-й неделе развития эмбриона(8 мм) можно видеть нервные волокна, идущие из крыловидной пластинки в краевой слой. По-видимому, это короткие волокна, которые, созревая, формируют межсегментные связи, обнаруживаемые в fasciculi proprii спинного мозга (система волокон, находящихся вблизи серого вещества спинного мозга (12) и связывающих между собой сегменты различных уровней). Вероятно, эти короткие волокна закладываются самыми первыми и находятся в непосредственной близости от клеток мантийного слоя. Логически можно предположить, что более длинные проекционные волокна, входящие в состав бокового канатика (funiculus lateralis, 11), должны образовываться в определенной последовательности, зависящей от их длины, и что, таким образом, возможно возникновение упорядоченной последовательности в развитии и расположении восходящих сенсорных волокон. Однако данных, подтверждающих существование такой последовательности, еще не получено. В то же время известно, что спинномозжечковые волокна, занимающие наиболее латеральное положение, появляются несколько позже, чем пучки, предшествующие им по времени формирования и лежащие более медиально. Волокна дорсального канатика (funiculus dorsalis, 13) в большинстве своем являются отростками клеток спинальных ганглиев (17). У эмбрионов на стадии 8 мм пучки этих волокон очень малы, на стадиях от 15 до 35 мм они быстро увеличиваются и уже у 35-миллиметровых эмбрионов они смещаются медиально, формируя настоящие задние (дорсальные) столбы спинного мозга. Существуют некоторые указания на то, что два пучка волокон, составляющих дорсальный канатик (fasciculus gracilis et cuneatus), формируются раздельно. На более поздних стадиях развития окончания волокон, идущих от нейронов спинальных ганглиев, видны среди группы нейронов дорсального мантийного слоя; эти волокна формируют другой пучок, лежащий дорсолатерально по отношению к указанной группе клеток, который, по всей вероятности, превращается в тракт, известный под названием тракта Лиссауэра.
Интенсивность деления клеток в базальной пластинке, очевидно, выше, чем в крыловидной, о чем свидетельствует тот факт, что спонтанные движения эмбриона появляются раньше, чем наступает момент, когда можно вызвать рефлекторные ответы на раздражение. Мотонейроны, иннервирующие скелетную мускулатуру, также созревают раньше нейронов высших отделов ЦНС (мозжечок, кора головного мозга). Общая скорость формирования нервных клеток, очевидно, одинакова в большей части спинного мозга и вполне достаточна для того, чтобы обеспечить избыточное число нейронов, необходимое для иннервации мышц туловища. Однако значительное число этих клеток в грудном отделе спинного мозга исчезает, оставляя скопления нейронов, иннервирующих определенные группы мышц. Остальные нейроны участвуют в формировании группы преганглионарных клеток интермедиолатеральной колонки (симпатический отдел вегетативной нервной системы).
Myelencephalon
Рис. 3.8. Myelencephalon, или продолговатый мозг (medulla oblongata), вначале формируется как структура, вполне сходная со спинным мозгом, и имеет крыловидную (4) и базальную (5) пластинки, крышу (6) и дно (7). Однако крыловидные и базальные пластинки раскрываются наподобие книжки, причем дно представляет как бы корешок. Происходит это в период образования мостового изгиба, когда крыша растягивается и становится очень тонкой. Клетки мезенхимы, прилежащие к крыше, дают начало большому числу гемоцитобластов и ангиобластов, образующих сосудистое сплетение, вдавливающее крышу; в конечном итоге из них образуется сосудистое сплетение четвертого желудочка. У эмбриона человека "зачаток" сосудистого сплетения можно видеть уже на 6-й неделе (12 мм), когда формирование клеток крови и сосудистой системы достаточно выражено. На стадии 22 мм видны эритробласты, окруженные выростами протоплазмы ангиобластов, которые превращаются в эндотелиальные клетки развивающегося сосудистого сплетения.

Рис. 3.8. Развитие продолговатого мозга. Стадии развития эмбриона:
А –10 мм; Б – 14,5 мм; В – 15 мм; Г – 73 мм. Показано боковое смещение крыловидной и базальной пластинок и истончение крыши, которая вдавливается формирующимся сосудистым сплетением. 1 – эпендимный слой; 2 – мантийный слой; 3 – краевой слой; 8 – пограничная бороздка; 9 – соматосенсорное ядро; 10 – соматомоторное ядро; 11 – висцеросенсорное ядро; 12 – висцеромоторное ядро; 13 – нижняя олива; з – клетки, идущие из крыловидной пластинки в "е"; и – клетки, идущие из базальной пластинки в "б"; к – клетки из эпендимы (1), идущие в оливарный комплекс (13). Остальные объяснения см. в тексте
Крыловидная и базальная пластинки (4.5) расщепляются в продольном направлении, так что из крыловидной пластинки образуются четыре клеточные колонки, а из базальной – три (рис. 3.8, Б – Г). Некоторые из черепномозговых нервов образуются из этих клеточных колонок или оканчиваются в них. Если начать с наиболее медиально расположенной колонки и идти в латеральном направлении, то ядрами, происходящими из них, будут:
а) п. hypoglossus– ядро подъязычного нерва (общесоматический эфферент);б) п. ambiguus – часть блуждающего нерва (специфический висцеральный эфферент) – клетки ядра этого нерва мигрируют в дальнейшем в вентролатеральном направлении; в) дорсальное ядро блуждающего нерва (общий висцеральный эфферент); г) дорсальное сенсорное ядро блуждающего нерва – п. tractus solitarius, (общий висцеральный афферент); д) сенсорное ядро языкоглоточного нерва (n. glossopharyngeus) – вкусовой нерв (специфический висцеральный афферент); е) нисходящая сенсорная часть ядра тройничного нерва (специфический соматический афферент); ж) ядро вестибулокохлеарного нерва(специфический соматический афферент).
Сенсорные ядра тройничного нерва (общий соматический афферент) и двигательные ядра тройничного и лицевого нервов (оба – специфические висцеральные эфференты) происходят из передней (ростральной) части продолговатого мозга; в задний мозг они перемещаются вторично. Это перемещение происходит после образования мостового изгиба в тот момент, когда каудальный отдел дна заднего мозга и ростральный отдел дна четвертого желудочка (IV) продолговатого мозга смыкаются друг с другом (рис. 3.5). Эти две области сливаются уже вторично, и создается впечатление, что некоторые структуры, происходящие из продолговатого мозга, будто бы сформировались в заднем мозге, хотя на самом деле их положение в заднем мозге взрослого обусловлено в первую очередь слиянием двух смежных областей дна первичного четвертого желудочка. К той же категории ядер относятся и ростральные отделы вестибулярных ядер (VIII). Закладка ядра отводящего нерва (VI) располагается каудальные ядра лицевого нерва (VII); оно занимает свое ростромедиальное положение благодаря сочетанию упомянутого выше слияния отделов продолговатого и заднего мозга и миграции его нейронов в медиальном направлении. Таким образом, выходит, что в действительности ни одно из ядер черепномозговых нервов (за исключением, возможно, небольшой части главного сенсорного и мезенцефалического ядер тройничного нерва) не образуется из клеток, первично формирующихся в заднем мозге, и конечное положение этих ядер в заднем мозге взрослого можно отнести за счет процессов их созревания и дифференцировки. В каудальных отделах мозга из крыловидной пластинки формируются ядра – п. gracilis, n. cuneatus и n. cuneatus accessorius, в которых оканчиваются волокна заднего (дорсального) канатика.
Рис. 3.9. Мост. На 7-й неделе (длина эмбриона около 16 мм) крыловидная пластинка дает начало клеткам, которые мигрируют в вентральном и ростральном направлениях из средней области продолговатого мозга и образуют на вентральной поверхности заднего мозга понтобульбарное тело и п. аrсиаtus (рис. 3.9, Б,Г). Понтобульбарное тело превратится в ядра моста (9), когда сверху к ним подойдут волокна из переднего мозга и, пройдя среди этих ядер, уйдут далее в спинной мозг. Образование нижней оливы и добавочной медиальной оливы начинается у эмбриона длиной 13 мм;принято считать, что они происходят из крыловидной пластинки продолговатого мозга. Между тем Бакстер <...> представил доказательства того, что эти ядра развиваются in situ посредством "конденсации" нейробластов, происходящих из базальной пластинки, и их пролиферации.

Рис. 3.9. Развитие области моста. Стадии развития эмбриона:
А – 13 мм; Б – 16мм; В – 100мм. Схематически представлена миграция клеток (а), которые образуют глубокие ядра мозжечка (б) и кору мозжечка (5). Г – миграция клеток, формирующих ядра основания моста из дорсальной поверхности продолговатого мозга в направлении от х к у. 1 – эпендимный слой крыши; 2 – крыловидная пластинка; 3 – базальная пластинка; 4 – пограничная бороздка; 7 – ретикулярные ядра; 8 – кортикоспинальные и кортикобульбарные волокна; 9 – ядро моста; 10 – вестибулокохлеарный нерв; IVv – четвертый желудочек. Остальные объяснения см. в тексте
Metencephalon
Метенцефалон, или задний мозг, можно разделить на три части: 1) центрально расположенную ядерную часть, которая служит прямым продолжением кпереди продолговатого мозга; 2) мозжечок и 3) мост, образованный волокнами и занимающий базальную область.
Рис. 3.9, В. Центральная часть заднего мозга – продолжение продолговатого мозга – также содержит крыловидную и базальную пластинки. Здесь происходит сходный с описанным выше процесс раскрытия, наподобие книжки, крыловидной пластинки, благодаря чему она сильно смещается в латеральном направлении. Вес это, конечно, сопровождается истончением крыши, но при этом на данном уровне не происходит вдавливания сосудистого сплетения. Вместо этого крыша разрастается вследствие миграции клеток из крыловидной пластинки и образует кору (5) и глубокие ядра (6) мозжечка. С крыловидной и базальной пластинками можно связать также формирование (б, в) различных ретикулярных ядер (7) заднего мозга.
Рис. 3.10. Мозжечок (cerebellum) частично развивается из ромбической губы (5), которая формируется в результате миграции клеток крыловидной пластинки в дорсальном направлении в момент, непосредственно предшествующий формированию мостового изгиба. Клетки ромбической губы располагаются на стыке крыши и крыловидной пластинки. При образовании мостового изгиба ромбические губы выпячиваются кнаружи (рис. 3.10, Б, В), создавая место образования в дальнейшем парных отверстий Люшка. Та часть ромбической губы, которая лежит ростральнее места образования этих отверстий, дает начало флоккулонодулярной дольке мозжечка(19, 20), тогда как клетки, расположенные каудальнее этих отверстий, примут участие в формировании вестибулярных ядер. Тело мозжечка (corpus cerebelli, 8), представляющее собой самый крупный отдел мозжечка у человека, развивается из клеток крыловидной пластинки ростральнее ромбических губ. Эти клетки мигрируют в дорсальном и медиальном направлениях и в дальнейшем, сливаясь, образуют червь (vermis, 16), расположенный по средней линии. После этого на каждой стороне червя быстро образуются выросты, формирующие передние и задние дольки (14, 15) полушарий мозжечка. Первые борозды мозжечка возникают в переднем отделе червя (16), затем они появляются на остальных отделах червя и лишь потом процесс формирования борозд медленно распространяется в латеральных направлениях по всей поверхности тела мозжечка.
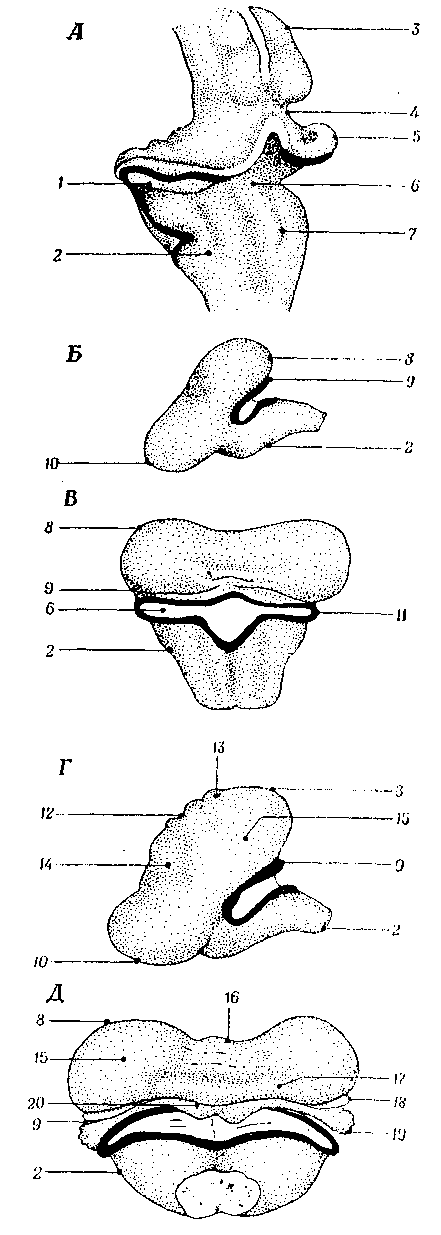
Рис. 3.10. Развитие мозжечка:
А – мозжечок и соседние области мозга (косая проекция стадия 28мм); Б, В и Г, Д – вид сбоку и вид сверху на стадиях 56 и 112 мм соответсвенно. А –- раскрытие крыловидных пластинок нa уровне мостового изгиба, при этом самые крайние части ромбической губы (5) становятся в дальнейшем тем местом, где образуется боковой карман четвертого желудочка и отверстие Люшка. 1 – закладка мозжечка; 2 – продолговатый мозг; 3 – крыша среднего мозга; 7 – пограничная бороздка; 9 – заднебоковая щель; 10 – мост; 11 – боковой карман четвертого желудочка; 12 – первая щель; 13 – посткливальная борозда; 14 – передняя доля; 15 – задняя доля; 17 – миндалина; 18 – paraflocculus. Остальные объяснения см. в тексте
Крыловидные пластинки заднего мозга распространяются рострально, покрывая четвертый желудочек (6), и сохраняются в "крыше" позади перешейка ромбовидного мозга (рис. 3.10, А, 4). В действительности эти выросты заднего мозга проходят кпереди за ростральную границу заднего мозга, смыкаясь с крышей среднего мозга.
Нижние ножки мозжечка (веревчатые тела) развиваются в центральной части крыловидной пластинки одновременно со спинальным трактом тройничного нерва и лежат кнаружи от волокон тройничного нерва в продолговатом мозге. Впервые их можно заметить в конце 2-го месяца развития, когда они представлены небольшим числом волоконец, направляющихся рострально в крыловидную пластинку заднего мозга и достигающих таким путем растущего мозжечка. Частично эти волокна начинаются от клеток, лежащих в основании задних рогов спинного мозга (в то время это еще крыловидная пластинка). В образовании веревчатых тел участвуют и другие волокна из нижней оливы и ретикулярных ядер продолговатого мозга. Верхняя ножка мозжечка(brасhiит conjuctivum) возникает в то же самое время, что и небольшие рассеянные волокна, идущие из развивающегося мозжечка в средний мозг. Эти волокна являются отростками клеток глубоких ядер мозжечка.
Isthmus rhombencephali
Рис 3.10, А. Эта область, представляющая собой сужение (4) формирующегося мозга, претерпевает множество изменений по мере роста крыловидной и базальной пластинок. В этой области из базальной пластинки развивается ядро блокового (IV) нерва (п. trochlearis– общий соматический эфферент). Нервные волокна из этих ядер идут в дорсальном направлении и на стадии 12 мм перекрещиваются в заднем мозге на уровне, лежащем каудально от перешейка. При последующем росте область базальной пластинки, включающая ядро блокового нерва, передвигается рострально, так что на стадии 26 мм это ядро лежит в среднем мозге, а перекрещивающиеся волокна теперь уже находятся в перешейке заднего мозга, где они располагаются и у взрослого. Ростральное смещение базальной пластинки в области перешейка заднего мозга на стадиях 12–16 мм сопровождается сходным смещением и крыловидной пластинки, вместе с которой перемещаются и некоторые ростральные отделы ядра тройничного нерва, в результате чего они располагаются кнаружи от образований базальной пластинки. Таким образом, мезенцефалический корешок тройничного нерва и образующие его клетки ложатся кнаружи от волокон блокового нерва.

Рис. 3.11. Развитие среднего мозга:
Стадии: А – 23 мм; Б – 56 мм; В – 112 мм. Показана миграция клеток из эпендимы (1) в ядро глазодвигательного нерва (5) и другие ядра среднего мозга, происхождение которых точно пока не установлено. Обратите внимание на то, что системы восходящих и нисходящих трактов связаны с вентральным отделом среднего мозга. 2 – краевая зона; 4 – дно; 6 – волокна, идущие из мозжечка; 7а – формирующиеся ядра бугорков четверохолмия; 8 – пограничная бороздка; 9 – мозжечковые волокна, окружающие красное ядро; 11 – медиальный продольный пучок. Остальные объяснения см. в тексте
Mescncephalon
Рис. 3.11. Этот отдел нервной трубки, также как и спинной мозг, имеет первичные крыловидную и базальную пластинки. Базальная пластинка (3) дает начало только одному большому ядерному скоплению (5) –ядру глазодвигательного (III) нерва (п. oculomotorius– общий соматический эфферент), которое хорошо выражено к концу 3-го месяца развития. Из крыловидной пластинки клетки (а) мигрируют в дорсальном направлении и образуют ядра нижних (7) и верхних (13) бугорков четверохолмия и, возможно, из нее некоторые клетки мигрируют вертикально и формируют красные ядра (в) (3-й месяц) наряду с отдельными ретикулярными ядрами (б). Детали формирования этих ядер еще до конца не известны; возможно, многие клетки могут образовываться in situ в результате пролиферации эпендимы и базальной пластинки. Еще менее ясно происхождение черного вещества (substantia nigra, г), которое не содержит пигмента ни у новорожденного, ни у 2–3-летних детей. В более поздний период развития на вентральной поверхности среднего мозга появляются два крупных скопления волокон (основания ножек мозга, basis pedunculi), которые начинаются в коре мозга и представляют собой двигательные пути, идущие к нижерасположенным центрам.
Рост крыловидной и базальной пластинок кнаружи сдавливает полость желудочка на уровне среднего мозга, сужая его до небольшого канала водопровода (aqueductus cerebri). Волокна восходящих медиальной и латеральной петель (10) проходят среди структур – производных крыловидной пластинки – и в течение третьего месяца распределяются по латеральному и вентральному отделам среднего мозга. Нисходящие кортикоспинальные волокна (12) приходят в средний мозг несколько позже, чем волокна петель, и накладываются на них с вентральной стороны.
Prosencephalon
Рис. 3.12 и 3.13. Наиболее ростральная часть нервной трубки, область плаща (pallium, 1), происходит из тех ее отделов, которые находятся кпереди от окончания sulcus limitans, и вторично делится на промежуточный мозг (diencephalon, 2) и конечный мозг (telencephalon, 3). Передний мозг развивается как вторичный ростральный вырост переднего конца нервной бороздки, которая в этот период открывается спереди отверстием (нейропором, 4). Открытый конец бороздки растет вперед и в завершение нейропор замыкается на ростральном конце выступающей части переднего мозга непосредственно перед тем, как эмбрион достигнет стадии 3 мм длины.
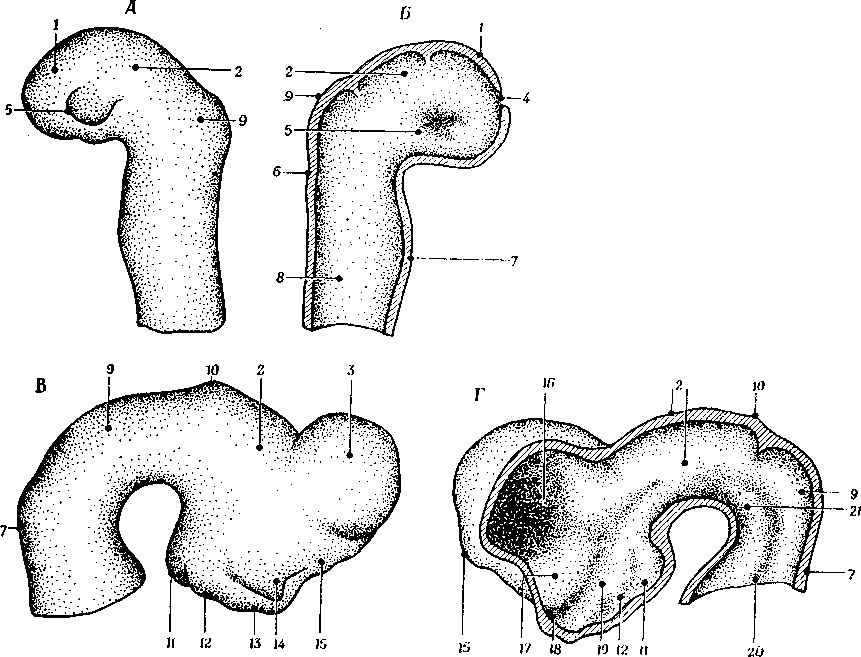
Рис. 3.12. Развитие переднего мозга и начальные стадии формирования конечного мозга:
А, Б – стадии 2,5 мм; В, Г – 10,2 мм; Б и Г – срединные сагиттальные разрезы; 6 – перешеек ромбовидного мозга; 7 – мост; 8 – продолговатый мозг; 9 – средний мозг; 10 – эпифиз; 11 – область сосцевидных тел; 12 – серый бугор; 13 – гипофизарный карман; 14 – глазной стебелек; 15 – обонятельная доля; 16 – межжелудочковое (монроево) отверстие; 17 – полосатое тело 18 – глазной карман; 19 – гипоталамус; 20 – пограничная бороздка; 21 – базальная пластинка. Остальные объяснения см. в тексте

Рис. 3.13. Развитие переднего мозга зародыша и формирование некоторых отделов промежуточного мозга (2), полосатого тела (17) и обонятельного мозга (15):
Стадии: А – 13,6 мм: Б – 60 мм: 6 – перешеек ромбовидного мозга: 14а – перекрест зрительных нервов; 16а – межжелудочковое отверстие; 19 – гипоталамус; 19а – гипоталамическая борозда; 20 – пограничная борозда; 22 – обонятельный мозг; 23 – борозда сосудистого сплетения; 24 – зрительный бугор; 25 – эпифиз; 26 – средний мозг; 27 – бугорки четверохолмия; 28 – гиппокампова борозда
В начальной стадии формирования передний мозг представлен коротким, слегка закругленным концом нервной трубки без каких-либо образований, сходных с полушариями (А и Б). Если передний мозг рассматривать сверху, то заметно, что он несколько сжат в ростральном отделе, тогда как в его задних отделах от той части, из которой развивается промежуточный мозг (2), в боковых направлениях отходят глазные пузыри. Изогнутая задненижняя часть промежуточного мозга несет на себе округлые выступы, которые латерально продолжаются в задненижнюю стенку глазного пузыря (5). Этот выступ превращается в перекрест зрительных нервов, идущих от обоих глаз. В данный период развития конечный мозг – это всего лишь нечетко ограниченный участок тонкой стенки рострального конца переднего мозга. Затем стенка первичного мозгового пузыря, соответствующая конечному мозгу, выпячивается в дорсолатеральном направлении и образует два мозговых пузыря, которые, разрастаясь и созревая, превращаются в полушария мозга (рис. 3.12, В и Г; см. также рис. 3.13). Полости этих пузырей образуют боковые желудочки полушарий. На ростральном конце нервной трубки и третьего желудочка находится участок паренхимы, соединяющий оба пузыря; этот участок в дальнейшем будет называться lamina terminalis. Крыша полушария очень тонка и в ростродорсальном направлении на небольшом протяжении образует передний мозг, а в каудальном направлении достигает крыши среднего мозга. Выросты конечного мозга образуются в результате усиленного деления клеток этой области; остающиеся отделы переднего мозга, расположенные каудальнее этих выростов, формируют промежуточный мозг (diencephalori).
Рис. 3.14. Вначале диенцефальный отдел развивается как массивное утолщение мантийного слоя (1), который разрастается так сильно, что краевого слоя фактически не остается (2). Существует некоторое различие в скорости деления клеток тех отделов, которые образуют дорсально расположенный таламус (24) и вентральный гипоталамус (19), в результате чего вдоль эпендимальной стенки формируется неглубокая бороздка (sulcus hypothalamicus) (19a). В передненижнем отделе промежуточного мозга образуются впячивания – глазные бокалы. Крыша промежуточного мозга становится крышей третьего желудочка, над которой лежит вдавливающее ее сосудистое сплетение.

Рис. 3.14. Развитие диенцефального отдела конечного мозга:
А – горизонтальный разрез; Б, В, Г – фронтальные разрезы; Б – 12 мм, разрез по линии х, показанной на рис. А; В – 17 мм; Г – 40 мм; Б – Г иллюстрируют развитие коры, полосатого тела (17) и промежуточного мозга (4) на 6-и, 7-й, 10-й неделях развития соответственно. Обращает на себя внимание рост нижнего отдела переднего мозга в направлении к промежуточному мозгу и ход нервных волокон вдоль плоскости соединения этих двух отделов (31). В конечном итоге эти пучки волокон образуют внутреннюю капсулу. 3 – эпендима; 5 – бахромка свода; 6 – концевая пластинка; 7 – мозговой пузырь; 8 – передний отдел таламуса; 9 – межжелудочковое отверстие; 10 – боковой желудочек; 11 – полушарие мозга; 13 – зрительный нерв; 14 – зрительный тракт; 18 – глазной карман; 19 – гипоталамус; 19а – гипоталамическая борозда; 28 – гиппокамп; 30 – чечевицеобразное ядро; IIIv – третий желудочек; а – клетки, мигрирующие из эпендимы; б – рост аксонов
Пузыри конечного мозга продолжают расти в ростральном, дорсальном и каудальном направлениях, покрывая собой промежуточный мозг (рис. 3.13 и 3.14). На ранних стадиях развития эти пузыри имеют очень тонкую стенку; в них заключено большое расширение центрального канала первичной нервной трубки. С ростом пузырей пластинка крыши сильно растягивается, а затем заворачивается в складку по мере того, как в дорсомедиальном направлении разрастаются полушария конечного мозга. Складка крыши образует сосудистую борозду (рис. 3.13, 23). Наружное капиллярное сплетение прилежит к крыше и, вдавливая ее в направлении, совпадающем с упомянутой выше складкой, образует сосудистое сплетение бокового желудочка (рис. 3.14,13а).
Таблица 3.1