

Биосинтез аминокислот
В атмосфере элементарный азот (Ν2) присутствует практически в неограниченном количестве. Прежде чем поступить в круговорот азота, он должен быть восстановлен до NH3 и включен («фиксирован») в аминокислоты.
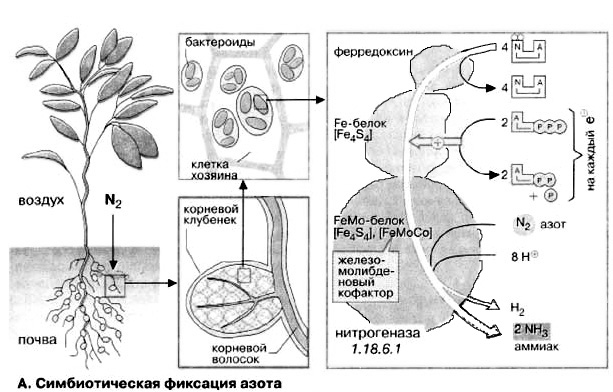
А. Симбиотическая фиксация азота
Фиксировать атмосферный азот способны лишь немногие виды бактерий и синезеленых водорослей. Они находятся в почве свободно или живут в симбиозе с растениями. Особо важное хозяйственное значение имеет симбиоз между бактериями рода Rhizobium и бобовыми растениями (Fabales), такими, как клевер, бобы или горох. Эти растения очень питательны благодаря высокому содержанию белка.
В симбиозе с бобовыми бактерии живут в корневых клубочках внутри растительных клеток, в так называемых бактероидах. С одной стороны, растение снабжает бактериоды питательными веществами, а с другой, извлекает пользу от фиксированного азота, который поставляет симбионт. Фиксирующим N2 ферментом бактерий является нитрогеназа. Она состоит из двух компонентов: Fe-белка и FeMo-белка. Fe-белок, содержащий [FeS4]-центр служит окислительно-восстановительной системой, которая принимает электроны от ферредоксина и передает их во второй компонент, FeMo-белок. Этот молибденсодержащий белок переносит электроны на N2 и таким образом через различные промежуточные стадии продуцирует NH3. Часть восстановительных эквивалентов переносится в побочной реакции на H+. Поэтому наряду с NH3 всегда образуется водород.
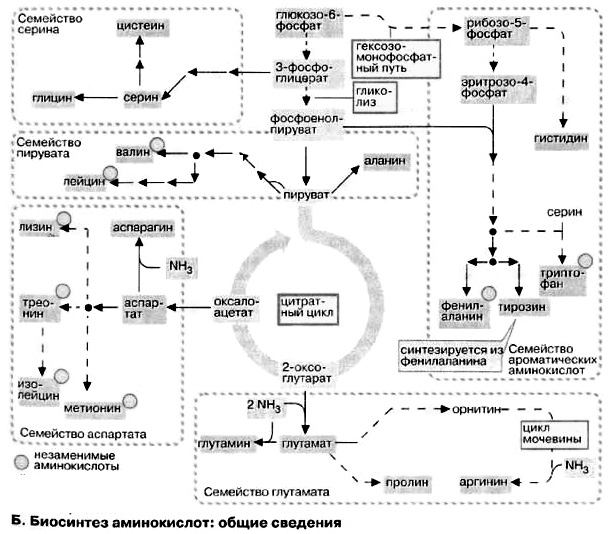
Б. Биосинтез аминокислот: общие сведения
По особенностям биосинтеза протеиногенные аминокислоты подразделяются на пять семейств. Члены каждого семейства имеют общих предшественников, которые образуются в цитратном цикле или при катаболизме углеводов.
В
то время как растения и микроорганизмы
вполне могут синтезировать все
аминокислоты, млекопитающие в ходе
эволюции утратили способность к синтезу
примерно половины из 20 протеиногенных
аминокислот. Поэтому незаменимые
аминокислоты
должны поступать с пищей. Так, организм
высших организмов не способен синтезировать
ароматические
аминокислоты
de novo (тирозин не является незаменимой
аминокислотой только потому, что может
образоваться из фенилаланина). К
незаменимым аминокислотам принадлежат
аминокислоты
с разветвленной боковой цепью: валин
и изолейцин, а также лейцин, треонин,
метионин и лизин. П
 итательная
ценность белков решающим образом
зависит от содержания незаменимых
аминокислот. Растительные белки зачастую
бедны лизином или метионином. В то же
время животных белки содержат все
аминокислоты в сбалансированных
соотношениях.
итательная
ценность белков решающим образом
зависит от содержания незаменимых
аминокислот. Растительные белки зачастую
бедны лизином или метионином. В то же
время животных белки содержат все
аминокислоты в сбалансированных
соотношениях.
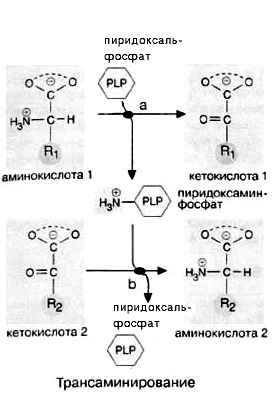
Заменимые аминокислоты (аланин, аспарагиновая и глутаминовая кислоты и их амиды, аспарагин и глутамин) образуются в результате трансаминирования из промежуточных метаболитов — 2-кетокислот. Пролин синтезируется в достаточных количествах из глутамата, а представители серинового семейства (серин, глицин и цистеин) сами являются естественными метаболитами организма животных.
Синтез белка
Генетический код
Большая часть генетической информации, содержащейся в ДНК, кодирует последовательность аминокислот. Процесс экспрессии генетической информации включает транскрипцию «текста», записанного на «языке нуклеиновой кислоты», в текст, записанный на «языке белков». Таково происхождение термина трансляция (дословно — перевод), используемого для обозначения процесса биосинтеза белков. Правила, которым следует трансляция, называют генетическим кодом.
Поскольку в биосинтезе участвуют 20 аминокислот, называемых протеиногенными, «язык» нуклеиновых кислот должен содержать по крайней мере 20 слов (кодонов) Однако в аминокислотном «алфавите» имеется только четыре «буквы» (А, Г, Ц и У или Т [или в англ. транскрипции: A, G, С и U или Т*]), так что для получения 20 различных слов каждое должно состоять по крайней мере из трех букв. Кодоны действительно включают три азотистых основания (триплет нуклеотидов). На схеме 1 представлен стандартный код ДНК (последовательность триплетов в некодирующей цепи), изображенный в виде круга. Схема читается от центра наружу, так что, например, триплет CAT кодирует аминокислоту гистидин. ДНК-кодоны идентичны таковым в мРНК (mRNA), за исключением того, что в мРНК вместо тимина (Т), характерного для ДНК, стоит урациял (U).
В качестве примера прочтения кода на схеме 2 показаны короткие участки нормального и мутантного гена β-глобина вместе с соответствующими последовательностями мРНК и аминокислот. Здесь показаны относительно часто встречающиеся точковые мутации, в результате которых остаток глутаминовой кислоты в положении 6 β-цепи заменен на валин. Такой мутантный гемоглобин в дезоксиформе склонен к агрегации, что вызывает деформацию эритроцитов и уменьшает эффективность транспорта кислорода (серповидноклеточная анемия).
В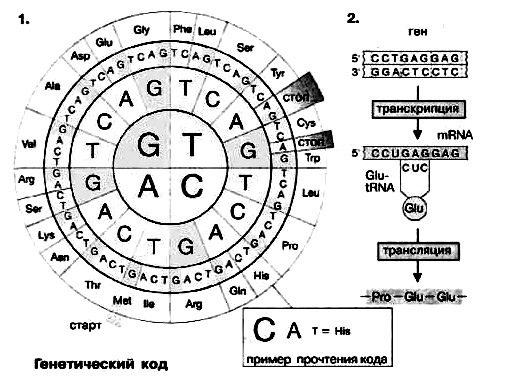 триплетном генетическом коде для 20
аминокислот потенциально существует
43
= 64 кодона. Таким образом, большинство
аминокислот записывается несколькими
кодонами, т. е. генетический код является
вырожденным.
Кроме того, имеются три триплета, которые
обозначают конец транскрипции
(стоп-кодоны).
Еще один специальный кодон, стартовый
(инициирующий) кодон, маркирует начало
трансляции. Генетический код, показанный
на рисунке, является почти универсальным.
Этому стандарту не полностью соответствуют
только митохондрии и некоторые
микроорганизмы.
триплетном генетическом коде для 20
аминокислот потенциально существует
43
= 64 кодона. Таким образом, большинство
аминокислот записывается несколькими
кодонами, т. е. генетический код является
вырожденным.
Кроме того, имеются три триплета, которые
обозначают конец транскрипции
(стоп-кодоны).
Еще один специальный кодон, стартовый
(инициирующий) кодон, маркирует начало
трансляции. Генетический код, показанный
на рисунке, является почти универсальным.
Этому стандарту не полностью соответствуют
только митохондрии и некоторые
микроорганизмы.