

Глава 5. Двигательные системы 115
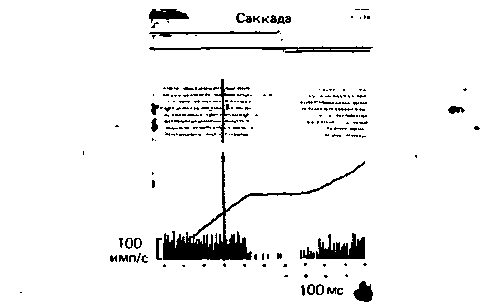
талыюму глазному полю. Следует упомянуть и о том, что аксоны нейронов сетчатой части черного вещества раздваиваются и одна из их ветвей идет к верхнему двухолмию среднего мозга; как известно, оно также связано с движениями глаз. На рис. 5.25 показана положительная корреляция между активностью этих нейронов и саккадами. Частота импульсации резко падает перед саккадой, что обусловлено тормозной стрионигральной связью. Такое отключение тормозного выхода черного вещества ведет к фазической активации следующего пункта-таламуса или верхнего двухолмия. О полном пространственном разделении скелетомоторной и окуломоторной петель свидетельствует, в частности, корреляция нейронной активности сетчатой части черного вещества с движениями либо глаз, либо рта, но никогда-с теми и другими одновременно.
Сложные петли (рис. 5.23, справа). К настоящему времени накоплены анатомические данные о существовании ряда «сложных» петель, начинающихся и заканчивающихся в лобных ассоциативных областях коры (дорсолатеральной префронтальной, латеральной орбитофронтальной, передней поясной), пройдя через ассоциативные ядра таламуса. В ходе филогенеза значительно возрастают размеры и значение корковых структур, стриатума и таламуса, участвующих в сложных петлях, так что у человека они становятся более обширными, чем двигательные петли. Однако функции сложных петель экспериментально не исследованы. Правда, регистрация активности индивидуальных клеток соответствующих участков коры, опыты с избирательным разрушением структур, а также клинические наблюдения на больных с поражениями лобных долей и лимбической системы свидетельствуют о том, что ассоциативные области коры ответственны за «высшие» (сложные) функции мозга. Как будет подробно рассматриваться в гл. 6 и 17, они играют определенную роль в формировании общего побуждения (драйва) и в регуляции конкретных драйвов, например голода, полового влечения. Они участвуют также в контроле мотивации, выборе стратегии и когнитивной деятельности. С другой стороны, все эти сложные функции могут нарушаться и у больных с поражениями базальных ганглиев. Следовательно, от базальных ганглиев принципиальным образом зависит проявление подобных функций и их включение в нормальную двигательную активность.
Связи между двигательными и сложными функциональными петлями базальных ганглиев. Если в пределах базальных ганглиев отдельные функциональные петли отделены друг от друга, возникает трудный вопрос, как и где происходит объединение когнитивных и мотивационных компонентов двигательной активности с элементами двигательных команд? В настоящее время любой ответ на
|
Рис. 5.25. Активность нейрона компактной части черного вещества обезьяны коррелирует с саккадами глаз. Вертикальной линией отмечен момент появления светового пятна. Обезьяна обучена саккадически переводить взгляд на эту новую точку фиксации. Примерно через 100 мс после появления светового пятна и за 200 мс до начала саккады нейронная активность почти полностью подавляется (перерыв точечной записи и гистограммы; плато на кумулятивной кривой). Это ведет к растормаживанию (активации) нейронов-мишеней в таламусе и крыше среднего мозга (0. Hikosaka, R. Η. Wurtz. J. Neurophysiol., 49. 1983) |
него будет умозрительным. Учитывая отсутствие связей между петлями в базальных ганглиях, можно, по крайней мере, утверждать, что они объединяются не здесь, а на уровне коры больших полушарий. Возможно, роль связующего звена принадлежит полю 6, которое, с одной стороны, является важнейшей мишенью двигательной петли, а с другой -получает входные сигналы от ассоциативной лобной коры.
Модуляция передачи информации в функциональных петлях базальных ганглиев. Прохождение информации в описанных выше множественных параллельных трансстриатных функциональных петлях может облегчаться или подавляться модулирующими системами. Эти механизмы представляют значительный интерес для клиники, поскольку в итоге функция базальных ганглиев определяется, очевидно, тонким балансом между ее облегчающей и тормозной модуляцией (см. ниже раздел «Патофизиология базальных ганглиев»). Уже продемонстрированы или предполагаются несколько модулирующих систем. Особого внимания среди них из-за своей роли в болезни Паркинсона заслуживает дофаминергическая. Дофаминергические интростриатные пути начинаются в сетчатой части черного вещества. Содержащие дофамин нейроны обнаружены также поодиночке или группами вне этого вещества, но только поблизости от него.
Очень тонкие дофаминергические аксоны сильно ветвятся, образуя по всему стриатуму относительно диффузную (т.е. без отчетливой топографической организации) сеть. Вдоль этих волокон находится множество крошечных, заметных в световой микроскоп утолщений.
116 ЧАСТЬ II. ДВИГАТЕЛЬНЫЕ И ИНТЕГРАТИВНЫЕ ФУНКЦИИ НЕРВНОЙ СИСТЕМЫ
называемых варикозностями. На электронных микрофотографиях они идентифицируются как пресинаптические элементы. У нейронов сетчатой части черного вещества довольно регулярная импулъсация с частотой около 1 Гц. Таким образом, каждую секунду импульс одной дофаминергической клетки вызывает высвобождение дофамина в многочисленных рассеянных по стриатуму синапсах.
Из-за своего диффузного строения дофаминергическая система непригодна для передачи детализированной, топографически организованной информации. Скорее ее можно рассматривать как своего рода «ирригационную систему», модулирующую передачу информации по главному каналу. Действительно, недавно было показано, что высвобождаемый в стриатуме дофамин модулирует глутаминергическую кортикостриатную передачу, хотя не вполне ясно, вызывает он торможение, облегчение или и то и другое. Восходящие дофаминергические волокна от среднего мозга направляются не только к стриатуму, но и к лимбическим структурам, и к префронтальной коре.
Аналогичное модулирующее влияние на базальные ганглии, возможно, оказывают серотоиинергические волокна от ядер шва, норадренергические волокна от голубого пятна, а также волокна с медиатором неизвестной природы от интраламинарных ядер таламуса и от миндалины; все они идут к стриатуму. Наконец, следует отметить, что в базальных ганглиях содержится множество местных нейронов (интернейронов), модулирующих поток информации в трансстриатных петлях. К ним относятся холинергические нейроны стриатума и различные, открытые лишь недавно пептидергическне нейроны.