

2. Первинна і вторинна будова днк
В природі роль генетичної нуклеїнової кислоти (гНК) виконують як ДНК (у більшості видів), так і РНК (деякі віруси). ДНК і РНК являють собою біополімери, мономерними ланками яких є залишки мононуклеотидів. Мононуклеотиди – це фосфорні ефіри нуклеотидів, кожен з яких в свою чергу побудований із залишка пентози (рибози або десоксирибози) і азотистої основи (похідної пурину або піримідину). До складу ДНК входять дві пуринових основи – аденін і гуанін – і дві піримідинових – цитозин і тимін. РНК замість тиміну містить урацил, а замість дезоксирибози – рибозу. Пентоза з’єднується з гетероциклічною основою N-глікозидним зв’язком. Таким чином, нуклеозидні залишки в ДНК і РНК відносяться до N-глікозидів.
Крім п’яти перерахованих нуклеотидів молекули РНК і ДНК містять у невеликих кількостях так звані «мінорні» нуклеотиди – дещо модифіковані та незвичайні (наприклад, метильовані) нуклеотиди.
Залишки нуклеотидів-мономерів у нуклеїнових кислотах з’єднані між собою фосфодиефірними зв’язками, які утворюються між 3'-вуглецевим атомом одного нуклеозидного залишку і 5'-атомом іншого. Тому зв’язок між двома сусідніми нуклеотидами називають 3'-5'-фосфодиефірним. Полінуклеотидні ланцюги ДНК і РНК полярні: на одному кінці завжди знаходиться вільна або заміщена група 3'-ОН, а на протилежному – фосфорильована або дефосфорильована група 5'-ОН. Ці кінці будь-якого полінуклеотиду позначають як 3' і 5'.
На початку 50-х років ХХ ст. видатним біохіміком Е.Чаргафом було встановлено, що кількість пуринових основ у молекулі ДНК завжди відповідає кількості піримідинових: А + Г = Ц + Т, вміст аденіну завжди дорівнює вмісту тиміну, а вміст гуаніну – вмісту цитозину:
А / Т = Г / Ц = 1.
Разом з тим відношення А + Т / Г + Ц є видоспецифічним і дуже відрізняється у різних видів.
Були одержані результати рентгеноструктурного аналізу ДНК М.Уілкінсон і Р.Франклін, які виявили високо впорядковану регулярну будову молекул ДНК.
Ґрунтуючись на цих дослідженнях, у 1953 р. Дж. Уотсон і Ф. Крік запропонували дволанцюгову модель вторинної структури ДНК. Згідно їх гіпотези ДНК складається із двох полінуклеотидних ланцюгів, закручених у праву спіраль один навколо другого і навколо спільної осі. Ці ланцюги є антипаралельними, утримуються разом водневими зв’язками між азотними основами, причому аденін завжди з’єднаний з тиміном, а гуанін з цитозином. Таким чином полінуклеотидні ланцюги є комплементарними. Спіраль закручена таким чином, що на її поверхні утворюється дві борозни: велика – шириною біля 2,20 нм і мала – шириною приблизно 1,20 нм. Діаметр спіралі дорівнює 1,80 нм, довжина витка – 3,40 нм, в одному витку спіралі вміщується 10 нуклеотидних залишків (рис.3). Пізніше було з’ясовано, що модель Уотсона і Кріка описує структуру однієї найбільш розповсюдженої подвійної спіралі, яка була названа В-формою або В-конформацією. Пізніше було встановлено існування інших форм ДНК, які можуть взаємно переходити одна в одну (табл.).
Таблиця.
Найважливіші конформації днк та деякі їх параметри
Конформація |
Довжина витка спіралі, нм |
Відстань між нуклеотидами, нм |
Кількість нуклеотидів на виток |
В А С Z |
3,40 2,80 3,10 – |
0,34 0,25 0,32 0,37 |
10 11 9 12 |
ДНК у В-формі спостерігається за високої відносної вологості (92%). Якщо остання складає 80%, то В-форма переходить у А-форму, для якої необхідна наявність іонів Na+, K+, Zs+. Це скорочує довжину ДНК приблизно на 25% у порівнянні з В-формою.
С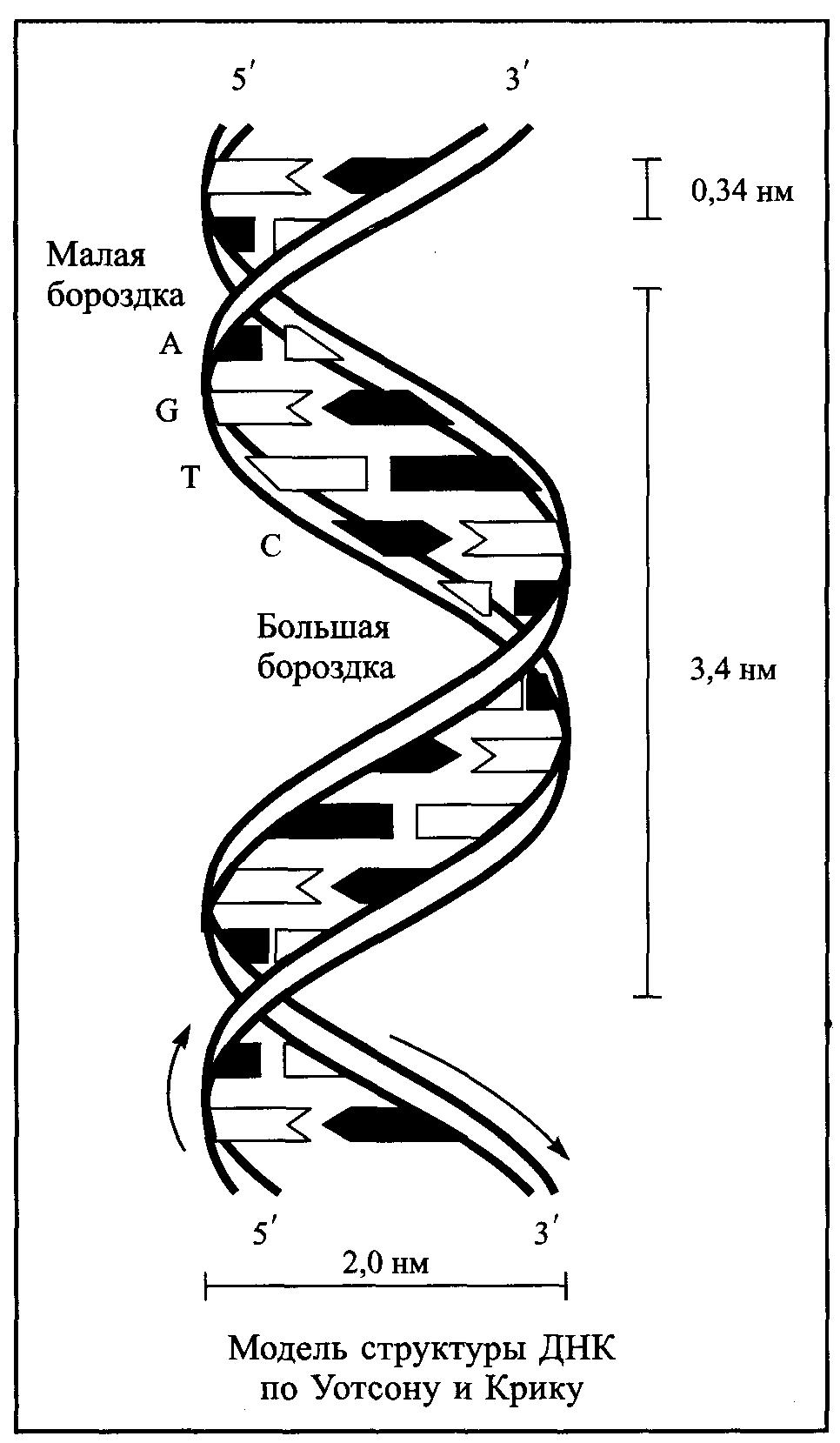 труктура
ДНК в А-формі характерна для гібридних
молекул ДНК-РНК, отже утворюється під
час синтезу РНК. За низької вологості
(66%) у присутності Li+
виникає С-форма. Вважають,
що С-форма частково утворюється у
хроматині, де ДНК асоційована з білками,
а також у складі деяких вірусів. Існують
ділянки ДНК, особливо багаті на Г – Ц
пари, які переходять у лівозакручену
Z-форму.
Z-форма має зигзагоподібний
вигляд, регулярна спіраль відсутня.
Вважають, що перехід право закрученої
форми до лівозакрученої форми слугує
сигналом, який контролює експресію
генів.
труктура
ДНК в А-формі характерна для гібридних
молекул ДНК-РНК, отже утворюється під
час синтезу РНК. За низької вологості
(66%) у присутності Li+
виникає С-форма. Вважають,
що С-форма частково утворюється у
хроматині, де ДНК асоційована з білками,
а також у складі деяких вірусів. Існують
ділянки ДНК, особливо багаті на Г – Ц
пари, які переходять у лівозакручену
Z-форму.
Z-форма має зигзагоподібний
вигляд, регулярна спіраль відсутня.
Вважають, що перехід право закрученої
форми до лівозакрученої форми слугує
сигналом, який контролює експресію
генів.
Рис 3. Модель структури ДНК за
Уотсоном і Кріком