

Взаємодія живого і косного компонентів
Хоча екосистема являє собою матеріально-функціональну цілісність, роль кожного компонента у визначенні властивостей цілого неоднакова. В зв’язку з цим однією з найважливіших характеристик екосистеми є кількісне співвідношення між масами живого і мертвого субстратів. Чим це відношення менше, тим, при інших рівних умовах, слабший вплив біоценозу на біотоп, менш ефективне кондеціонування середовища, напрямлене на посилення гомеостазу системи. Як правило, косний компонент у водних екосистемах представлений значно сильніше, ніж в наземних. Середня біомаса організмів на суші досягає 10 кг/м2, а в гідросфері – близько 10 г/м2. А якщо ще врахувати, що область поширення життя на суші вимірюється по вертикалі десятками метрів, а у воді переважно сотнями метрів і кілометрами, то маленька концентрація життя в гідросфері стає ще очевиднішою. Але чим менша водойма, тим вища в ній концентрація живого. Проте, живому компоненту належить активна і провідна роль в біогеоценотичних процесах. Пристосовуючись до абіотичного оточення, біоценоз безперервно еволюціонує як система.
Хоча ідеальна схема коло обігу речовин передбачає їх переміщення по замкненому колу, в реальних екосистемах спостерігається інша картина. Наприклад, кисень, який виділяється фото синтезуючими організмами, в певній мірі використовується на дихання тварин, але частина його тимчасово виходить з коло обігу, переміщуючись в атмосферу. Органічна речовина після відмирання організму частково опускається на грунт і захоронюється в товщі донних осадів.
Сам процес взаємодії косного і живого компонентів екосистеми далеко не завжди відповідає уявленню про колообіг речовин. Тільки по відношенню до Світового океану в цілому можна говорити про практично реалізований принцип циркуляції речовин по замкненому колу, хоча навіть і тут є резервати.
В стічних озерах процес виходу речовин з коло обігу в атмосферу та відкладення його на дні доповнюється винесенням як мертвого матеріалу, так і живих організмів (біосток). Ще сильніше виражені відхилення від ідеальної схеми у водосховищах та ріках, до яких застосовують термін «транзитний», або «поточний» коло обіг. Організми, котрі народжуються у верхній частині ріки, зносяться вниз течією і більше не беруть участі в процесах коло обігу в місці свого виникнення.
Стійкість
екосистем
Це
поняття відрізняється певною
невизначеністю, оскільки в нього
вкладається найрізноманітніший зміст.
Перш за все, стійкою може називатися
система, котра відрізняється постійністю
свого стану, відсутністю змін. Другий
критерій – інерція, стійкість до
порушень, третій – «еластичність»,
здатність повертатися до вихідного
стану після порушення зовнішніми силами.
Четвертий критерій – амплітуда змін у
відповідь на зовнішню дію, величина
відхилення, при якому система може
повертатися у вихідний стан. Особливою
формою стійкості системи є циклічність
змін, наявність стабільних лімітованих
циклів. Стійкість структурних (Sс)
і функціональних (Sф)
характеристик екосистем можна оцінити
середнім значенням відхилень вимірюваної
величини відносно середньої(). Як міру
гомеостазу передбачається використовувати
величину G1=1
-
 при Sс
≥
Sф.
при Sс
≥
Sф.
Система рахується тим стабільнішою, чим менше стан одних компонентів (функцій) залежить від коливання інших. По деяких розрахунках стабільність планктонних систем в 6-7 разів вища, ніж в оточуючого їх комплексу абіотичних факторів.
Велике число можливих поведінкових реакцій посилює стабільність (стійкість) живих систем в умовах різких локальних і тимчасових змін в абіотичному середовищі. Гомеостатичні механізми екосистем являють собою буфер, який згладжує амплітуду змін систем в певних межах.
Стійкість живих систем також переважно зростає з ускладненням їх структури, особливо видової і трофічної, що в свою чергу супроводжується характерними зрушеннями в енергетиці угрупувань.
В якості
прикладу можна навести результати
гідробіологічних дослідів, виконаних
на Чорному морі та в тропічній частині
Тихого океану. Для характеристики
складності трофічної структури
використовувався символ f;
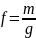 де m
– число харчових зв’язків в піддослідних
планктонних угрупуваннях, а g
– число екологічних груп в них. В
планктонній популяції Чорного моря
кількість екологічних груп становила
14, трофічних зв’язків – 50, відповідно
індекс f
становив 3.6; в планктонній популяції
тропічної частини Тихого океану к-ть
екологічних груп становила 14, трофічних
зв’язків – 50, а індекс f
становив 7.1. Тобто, в Чорному морі індекс
трофічної складності був майже в два
рази меншим, і, відповідно, система була
менш стійкою. В стійких популяціях
спостерігається відносно постійний (в
середньому 27 – 28%) показник ефективності
передачі речовини і енергії через всі
трофічні рівні. В малостійких угрупуваннях
інтенсивність потоку з переходом на
кожний наступний трофічний рівень і
затрата енергії зростають, ефективність
передачі енергії різко зменшується.
де m
– число харчових зв’язків в піддослідних
планктонних угрупуваннях, а g
– число екологічних груп в них. В
планктонній популяції Чорного моря
кількість екологічних груп становила
14, трофічних зв’язків – 50, відповідно
індекс f
становив 3.6; в планктонній популяції
тропічної частини Тихого океану к-ть
екологічних груп становила 14, трофічних
зв’язків – 50, а індекс f
становив 7.1. Тобто, в Чорному морі індекс
трофічної складності був майже в два
рази меншим, і, відповідно, система була
менш стійкою. В стійких популяціях
спостерігається відносно постійний (в
середньому 27 – 28%) показник ефективності
передачі речовини і енергії через всі
трофічні рівні. В малостійких угрупуваннях
інтенсивність потоку з переходом на
кожний наступний трофічний рівень і
затрата енергії зростають, ефективність
передачі енергії різко зменшується.