

216 Глава 9
один нейрон, В, причем синапс А->С слаб, а синапс В-^С весьма эффективен; допустим далее, что А и В разряжаются почти одновременно или же В чуть раньше А и что С дает разряд не под влиянием А, а в результате сильного воздействия В. В модели Хебба уже сам факт, что С разряжается сразу после А, повышает эффективность синапса А—*С. Мы предположим также, что если импульсы, приходящие к С от А, не сопровождаются импульсами в С, то синапс А-^С ослабевает.
Применим эту модель к бинокулярной конвергенции у нормального животного. Пусть клетка С — бинокулярная, аксон А идет от не доминантного, а аксон В — от доминантного глаза. Недоминантный глаз будет реже активировать клетку С, чем доминантный. Согласно гипотезе Хебба, синапс А-^С будет сохраняться или усиливаться, пока за импульсами в А будут следовать импульсы в С, а такая последовательность более вероятна, если в надлежащий момент времени от другого глаза по аксону В постоянно поступает «подкрепление». А именно это и будет происходить при точном совмещении изображений на обеих сетчатках. Если активность в А не сопровождается активностью в С, синапс А-^С за длительный период времени ослабеет. Получить прямое доказательство, что модель Хебба применима к косоглазию, — дело нелегкое (по крайней мере в ближайшем будущем), но сама идея, мне кажется, заслуживает внимания.
Анатомические последствия депривации
То, что в коленчатых телах, где нет или почти нет возможностей для конкуренции глаз, мы не смогли обнаружить заметных физиологических дефектов, казалось, подтверждает идею о том, что последствия закрытия одного глаза связаны с конкуренцией, а не с бездействием глаза. Правда, клетки коленчатого тела были атрофичными, но — рассуждали мы — вряд ли можно ожидать, что все будет ясно. Если конкуренция и в самом деле играет важную роль, то корковый слой 4С мог, как нам казалось, быть подходящим объектом для проверки этой идеи, поскольку клетки в нем тоже монокулярны и конкуренция маловероятна, так что чередование полосок, связанных с левым и с правым глазом, должно остаться ненарушенным. Итак, путем длинных микроэлектродных проходок через слой 4С мы начали выяснять, сохраняются ли такие полоски после закрытия одного глаза и остаются ли нормальными их размеры. Вскоре стало ясно, что слой 4С по-прежнему подразделяется на участки левого и правого глаз, как у нормальных животных, и что клетки в полосках, соединенных с ранее закрытым глазом, остались практически нормальными. Однако последовательности клеток с доминированием закрытого глаза оказались очень короткими, как если бы полоски были аномально узкими (около 0,2 мм вместо 0,4 или 0,5 мм), а полоски, принадлежащие открытому глазу, — соответственно более широкими.
Как только это стало доступным, мы начали использовать методику с инъекцией метки в глаз и транснейронным переносом ее в кору, чтобы получить прямое и наглядное подтверждение сделанных выводов. По прошествии периода депривации длительностью в несколько месяцев мы вводили кошке или обезьяне в тот или в другой глаз радиоактивную аминокислоту. Радиоавтографы показали заметное сужение полосок, связанных с ранее депривирован-ным глазом, и соответственное расширение полосок, принадлежащих нормальному глазу. Микрофотография слева на рис. 145 иллюстрирует результаты инъекции радиоактивной аминокислоты в нормальный глаз. На этом снимке, сделанном, как обычно, в условиях темного поля, представлен срез, параллель-

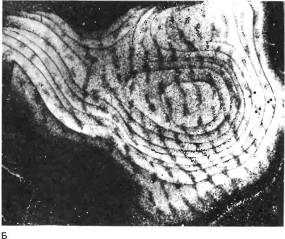
Рис. 145. Срезы зрительной коры макака, правый глаз которого был закрыт с момента рождения до 18-месячного возраста. В левый глаз инъецировали радиоактивную аминокислоту, и неделю спустя приготовили срезы мозга, параллельные поверхности зрительной коры. Кора имеет куполообразную форму, поэтому такие срезы вначале идут тангенциально, а затем образуют кольца (Л) все большего диаметра, подобные кольцам луковицы. Эти кольца были вырезаны из микрофотографий и склеены вместе (Б) . Впоследствии мы научились расправлять кору перед ее замораживанием, и тогда уже не приходилось разрезать и склеивать фотографии серийных срезов. На обычном радиоавтографе зерна серебра выглядят черными на белом фоне. Мы, однако, применяли метод исследования в темном поле, при котором зерна серебра рассеивают свет и их скопления выглядят как светлые участки. Светлые полоски — места поступления в слой 4С меченых атомов из открытого глаза — расширены, а темные (связанные с закрытым глазом) сильно сужены.