

Глава 9
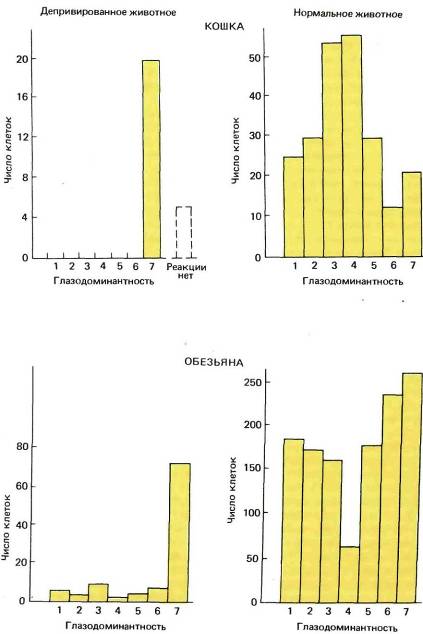
Рис. 133. Правый глаз котенка был на два с половиной месяца лишен зрительных стимулов в возрасте примерно десяти дней, когда глаза обычно уже открыты. В этом эксперименте (верхние гистограммы) мы зарегистрировали ответы только двадцати пяти клеток. (В последующих экспериментах мы смогли исследовать больше клеток и обнаружили, что небольшая доля их испытывала влияние со стороны ранее закрытого глаза.) В опытах на детеныше обезьяны (нижние гистограммы) результаты оказались весьма сходными. Его правый глаз закрыли в возрасте двух недель и держали закрытым на протяжении 18 месяцев. Позднее мы нашли, что те же результаты получаются и при закрытии глаза всего на несколько недель.
ДЕПРИВАЦИЯ И РАЗВИТИЕ 203
Для того чтобы определить локализацию дефекта, нужно было, очевидно, регистрировать реакции от более низких уровней, начиная, скажем, с сетчатки или с коленчатого тела. Результаты оказались однозначными: как в сетчатке, так и в коленчатом теле было множество клеток с практически нормальными реакциями. Клетки тех слоев коленчатого тела, к которым шли волокна от ранее закрытого глаза, имели обычные рецептивные поля с центром и периферией; они хорошо реагировали на маленькие пятна и плохо — на рассеянный свет. Единственным намеком на аномалию была некоторая вялось реакций этих клеток по сравнению с клетками слоев, получавших входные сигналы от нормального глаза.
Зная об этой относительной нормальности, мы были поражены, впервые увидев под микроскопом наружное коленчатое тело, окрашенное по Нисслю. Его аномальность была видна практически и без микроскопа. Коленчатое тело у кошки организовано несколько проще, чем у обезьяны; оно состоит в основном из двух крупноклеточных слоев, расположенных наверху, а не внизу, как у обезьяны. Верхний слой имеет вход от контралатерального, а нижний — от ипсилатерального глаза. Под этими слоями лежит довольно плохо выраженный слой мелких клеток с несколькими подразделениями, которые я здесь игнорирую. На каждой стороне крупноклеточный слой, получавший входы от закрытого глаза, был бледнее и явно тоньше своего партнера, который выглядел совершенно нормальным. Клетки аномальных слоев были не только бледными, но и сморщенными примерно до двух третей своего нормального поперечного сечения. Результат, полученный при закрытии правого глаза, показан на рис. 134. Сходные результаты были получены и на макаках (рис. 135). Таким образом, мы столкнулись с парадоксом, на разрешение которого потребовалось несколько лет: в клетках наружного коленчатого тела, физиологически казавшихся сравнительно нормальными, гистологическое исследование показывало явную патологию. Как бы то ни было, мы теперь имели ответ на наш исходный вопрос, поскольку корковые клетки, практически не отвечавшие на стимуляцию ранее закрытого глаза, тем не менее, видимо, получали значительный и как будто бы нормальный поток входных сигналов от коленчатого тела; это позволяло думать, что первичное повреждение локализуется не в глазу или коленчатом теле, а главным образом в коре. При гистологическом исследовании коры мы не находили абсолютно никаких признаков аномалии. Как мы увидим далее, в коре все же были анатомические дефекты, но они не выявлялись с помощью применявшихся нами методов окраски.
Следующий вопрос заключался в том, что же именно порождает аномалию при закрытии глаза. Сшивание век уменьшает количество света, достигающего сетчатки, приблизительно в 10—50 раз; оно, разумеется, препятствует также созданию на сетчатке каких-либо изображений. Может ли аномалия вызываться просто уменьшением количества света? Чтобы выяснить это, мы вставляли в один глаз новорожденного котенка матовую контактную линзу из пластмассы, по консистенции близкой к материалу мячика для настольного тенниса. У других животных мы вместо этого пришивали на одном глазу тонкую просвечивающую мембрану, играющую роль дополнительного века и называемую мигательной перепонкой, которая имеется у кошек, но отсутствует у людей. Пластмасса или перепонка уменьшала количество света всего лишь вдвое, но предотвращала формирование каких-либо сфокусированных изображений. Результаты оказались такими же: аномальная корковая физиология и аномальная гистология коленчатого тела. Очевидно, повреждение вызывалось невозможностью воспринимать формы, а не просто недостатком света.
У нескольких котят перед регистрацией нейронных ответов мы тестировали