

184 Глава 8
при включении всех трех проекторов. Выберем теперь другой участок, например оранжевый, и отрегулируем поочередно интенсивность каждого проектора так, чтобы показания фотометра для оранжевого участка совпадали с теми, которые мы получили ранее для зеленого. Таким образом, при включении трех проекторов свет, приходящий теперь от оранжевого участка, идентичен по составу тому, который минутой ранее приходил от зеленого участка. Что мы ожидаем увидеть? Рассуждая примитивным образом, мы скажем, что оранжевый участок превратился в зеленый. Но он все-таки выглядит оранжевым — его цвет даже нисколько не изменился. Мы можем повторить этот эксперимент с любыми двумя участками. Вывод таков: не столь важно, на какую интенсивность света установлены три проектора, пока от каждого из них поступает какой-то свет. Почти в любом случае мы заметим лишь очень малые изменения воспринимаемого цвета.
Такие эксперименты убедительно показали, что ощущение, возникающее в какой-либо части поля зрения, зависит как от света, приходящего от этой части, так и от света, приходящего от остальных участков. Иначе как мог бы свет одинакового спектрального состава вызывать в одном случае ощущение зеленого, а в другом — ощущение оранжевого? Принцип, применимый к черному, белому и серому и столь четко сформулированный Герингом, оказывается верным и в отношении цвета. Для цвета мы имеем оппонентность не только локальную (красный/зеленый и желтый/синий), но также и пространственную: красный/зеленый в центре против красного/зеленого на периферии и аналогичную оппонентность для желтого/синего.
В 1985 году Дэвиду Инглу в лаборатории Лэнда удалось в аквариуме с подводной мозаикой из разноцветных прямоугольников обучить золотую рыбку подплывать к участку определенного цвета. Он обнаружил, что рыбка плывет к одному и тому же цвету, например синему, независимо от спектрального состава света: она, как и мы, выбирает синий участок, даже если свет от него идентичен по составу свету, который в предыдущей пробе при ином источнике света приходил от желтого лоскута, отвергаемого рыбкой. Таким образом, рыба тоже выбирает участок поверхности по его собственному цвету, а не по спектральному составу отражаемого им света. Это означает, что феномен константности цвета не следует считать каким-то усовершенствованием, недавно добавленным в ходе эволюции к цветоощущению некоторых высших млекопитающих, в том числе и человека; наличие его у рыб указывает на то, что это примитивный, весьма общий аспект цветового зрения. Очень заманчиво (и довольно несложно) было бы проверить, обладают ли той же способностью насекомые с цветовым зрением. Я думаю, что это именно так.
Лэнд и его группа (среди прочих Дж. Мак-Кэнн, Н. Доу, М. Берне и X. Перри) разработали несколько процедур для предсказания видимого цвета некоторого объекта по спектрально-энергетическому составу света от всех точек поля зрения, но без какой-либо информации о источнике света. Расчет заключается в том, что для каждого из трех отдельных проекторов определяют отношение света, приходящего от места, цвет которого нужно предсказать, к усредненному свету, приходящему от окружения. (Площадь «окружения», которую нужно учитывать, в разных вариантах теории Лэнда различна. В новейшем варианте предполагается, что влияние окружающих участков уменьшается с расстоянием.) Получаемая тройка чисел — отношения берутся для каждого проектора — однозначно определяет цвет данного места. Любому цвету, таким образом, можно поставить в соответствие некоторую точку в трехмерном пространстве, координатными осями которого будут три отношения, полученные для красного, зеленого и синего света. Чтобы сделать формулировку как можно более реалистичной, три источника света подбираются в соответствии с
ЦВЕТОВОЕ ЗРЕНИЕ 185

Рис. 124. В экспериментах Дэвида Ингла золотая рыбка обучалась подплывать за вознаграждение — кусочек печенки — к полоске заданного цвета. Она плывет, например, к желтой полоске независимо от соотношения интенсивностей света от трех проекторов. Такое поведение поразительно сходно с константностью восприятия цвета у человека.
кривыми спектральной чувствительности трех типов колбочек человека.
То, что таким образом можно вычислить цвет, означает наличие константности цвета, поскольку для каждого проектора подсчитывается соотношение света от одной области к свету от усредненного окружения. Точная установка силы света в проекторах более не важна: единственным условием остается то, что мы должны иметь какой-то свет от каждого проектора — иначе нельзя будет вычислить никакого отношения. Одно из следствий всего этого заключается в том, что для появления цвета необходимы различия в спектральном составе света в пределах поля зрения. Для восприятия цвета нам нужны цветовые границы, точно так же как нужны границы освещенности для восприятия черного и белого. Вы можете легко убедиться в этом, снова использовав два диапроектора. Поставьте перед одним из проекторов красный фильтр (подойдет красный целлофан) и осветите любую группу предметов. Я предпочитаю брать белую или желтую рубашку и яркий красный галстук. При таком освещении ни рубашка, ни галстук не выглядят вполне красными: и то и другое кажется розоватым и как бы полинявшим. Теперь осветите ту же комбинацию предметов вторым проектором, прикрыв его синим целлофаном. Рубашка будет выглядеть бледно-голубоватой, а галстук — черным: красные объекты не отражают коротких световых волн. Вернитесь к красному проектору и убедитесь еще раз, что галстук не выглядит особенно красным. Теперь добавьте синий проектор. Вы знаете, что при добавлении синего света вы не получите от галстука какого-либо дополнительного отражения — вы только что это проде-
186 ГЛАВА 8
монстрировали, — но при включении синего проектора красный галстук внезапно засверкает хорошим ярким красным цветом. Это убедит вас в том, что галстук делают красным не только лучи, приходящие от него самого.
Эксперименты со стабилизированными цветовыми границами согласуются с представлением о том, что вообще для видения цвета необходимы различия на границах. Альфред Ярбус, имя которого упоминалось в главе 4 в связи с движением глаз, в 1962 году показал, что если смотреть на синее пятно, окруженное красным фоном, стабилизация границы пятна на сетчатке приводит к его исчезновению: синее пропадает, и все, что можно видеть, — это красный фон. Стабилизация границ на сетчатке явно делает их неэффективными, а без них исчезает и цвет.
Эти психофизические доказательства того, что для восприятия цвета нужны различия в спектральном составе света от разных участков поля зрения, указывают на возможное наличие в наших сетчатках или мозгу клеток, чувствительных к цветовым границам. Этот довод аналогичен тому, который приводился нами в главе 4 по поводу восприятия черных или белых объектов. Если на каком-то уровне нашей зрительной системы информация о цвете передается только на линиях цветового контраста, то клетки с рецептивными полями, лежащими целиком внутри областей однородного цвета, будут бездействовать. Результат — экономия при обработке информации. Таким образом, от передачи сведений о цвете лишь на границах мы получаем два преимущества: во-первых, цвет не меняется при изменениях освещения, так что мы узнаём о свойствах рассматриваемых объектов без искажений, вносимых источником света;, во-вторых, информация обрабатывается экономным способом. Теперь мы можем задаться вопросом: почему система эволюционировала именно таким образом? Была ли главным фактором эволюции необходимость константности цвета, а экономия:— лишь сопутствующей выгодой? Или, наоборот, экономия играла ведущую роль, а константность — второстепенную. Второе предположение многим может показаться более убедительным: вряд ли эволюция могла предвидеть появление ламп накаливания или флуоресцентного освещения, да и наши рубашки вовсе не были такими белыми, пока не появились современные моющие средства.
Физиология цветового зрения: ранние результаты
Первая физиологическая информация на клеточном уровне была получена спустя 250 лет после Ньютона в исследованиях шведско-финско-венесуэль-ского физиолога Гуннара Светихина, который в 1956 году на костистой рыбе осуществил внутриклеточную регистрацию активности нейтронов сетчатки — сначала он принял их за колбочки, но они оказались горизонтальными клетками. На освещение сетчатки эти клетки отвечали только медленными потенциалами (потенциалов действия не наблюдалось). Как показано на рис. 125, Свети-хин обнаружил три типа клеток: первый тип, названный им L-клетками, гиперполяризовался при световой стимуляции независимо от спектрального состава света; второй тип, названной r-g-клетками, гиперполяризовался волнами малой длины с максимумом ответа на зеленый свет и деполяризовался волнами большой длины с максимумом ответа на красный свет; третий тип, названный с учетом теории Геринга у-клетками, отвечал по типу клеток r-g, но с максимумом гиперполяризации на синий и максимумом деполяризации на желтый свет. У клеток r-g и у-Ь белый свет вызывал лишь слабые и быстро затухающие ответы, как и следовало ожидать ввиду широкополосного спектрально-энергетического состава белого света. Кроме того, у клеток обоих этих типов, которые мы можем назвать оппонентно-цветовыми клетками, свет с некоторой
ЦВЕТОВОЕ ЗРЕНИЕ 187
Рис. 125. Гуннар Светихин и Эдуард Мак-Никол регистрировали реакции горизонтальных клеток на цвет у костистой рыбы. Отклонения вниз от серой линии соответствуют гиперполяризации, а отклонения вверх — деполяризации.
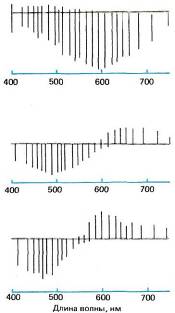
промежуточной длиной волны, называющейся точкой перекреста, не вызывал никакой реакции. Поскольку эти клетки реагируют на окрашенный, но не на белый свет, они, вероятно, связаны с цветовыми ощущениями.
В 1958 году Рассел де Валуа и его сотрудники зарегистрировали ответы, поразительно сходные со светихинскими, от клеток наружного коленчатого тела макака. Ранее с помощью поведенческих тестов де Валуа показал, что цветовое зрение макака и человека почти одинаково; например, соотношение, в котором нужно смешать два цветных луча, чтобы получить третий цвет, почти идентично у обоих видов. Поэтому можно думать, что макаки и люди обладают сходными механизмами на нижних уровнях зрительной системы, и мы, по-видимому, вправе сравнивать психофизику цвета у человека с физиологией у макака. Де Валуа обнаружил, что многие клетки коленчатого тела активировались рассеянным монохроматическим светом в диапазоне от одного конца спектра до точки перекреста, где реакция отсутствовала, и подавлялись светом во втором диапазоне — от точки перекреста до другого конца спектра. И вновь аналогия с цветовыми процессами Геринга была полной: де Валуа выявил ошюнентно-цветовые клетки двух типов, красно-зеленые и желто-синие; у каждого типа смешение двух световых волн, длины которых на волновой шкале располагались симметрично относительно точки перекреста, приводило к взаимному погашению реакций точно так же, как в восприятии добавление синего к желтому или зеленого к красному порождает белый. Результаты де Валуа особенно хорошо согласовались с формулировками Геринга, поскольку две группы цветовых клеток имели максимумы реакции и точки перекреста точно в тех местах спектральной шкалы, чтобы одна группа могла отражать «желто-синие» свойства падающего света, а другая группа — «красно-зеленые».