

112 Глава 5
свойственны простые рецептивные поля и ориентационная чувствительность. Если перейти к следующему уровню — к слоям, лежащим выше и ниже слоя 4С, — то подавляющее большинство клеток окажутся сложными. Клетки, реагирующие на концы линий, в слоях 2 и 3 составляют около 20 процентов, а в других слоях встречаются редко. Таким образом, в целом обнаруживается явная корреляция между сложностью клеток и положением их в зрительном пути, которое можно оценить по числу синаптических переключений до данного места.
В утверждении, что большинство клеток выше и ниже слоя 4 — сложные, упускаются из виду существенные различия между слоями: сложные клетки здесь далеко не одинаковы. Разумеется, все они имеют одно общее свойство, характерное для сложных клеток, — на движущуюся линию оптимальной ориентации они отвечают по всему рецептивному полю, независимо от конкретного положения стимула. Однако они различаются между собой другими свойствами. Можно выделить четыре подтипа клеток, которые содержатся в основном в разных слоях. Большинство сложных клеток в слоях 2 и 3 отвечает тем лучше, чем длиннее стимульная линия (в этом проявляется свойство суммации по длине). Однако реакция таких клеток становится слабее, когда длина стимула превышает некоторую критическую величину (если данная сложная клетка относится к нейронам, реагирующим на концы линий). Что касается клеток слоя 5, то для них короткие линии, занимающие по длине лишь небольшую долю рецептивного поля, почти так же эффективны, как и длинные; рецептивные поля этих клеток гораздо больше, чем у клеток в слоях 2 и 3. Напротив, клетки слоя 6 реагируют тем лучше, чем длиннее линия с оптимальной ориентацией (их ответ начинает ухудшаться только тогда, когда линия займет всю длину рецептивного поля, которая в несколько раз больше его ширины, т. е. рецептивное поле длинное и узкое). Можно сделать вывод, что аксоны, выходящие из слоев 5 и 6 и из слоев 2 и 3 и идущие к разным участкам мозга (к верхним бугоркам четверохолмия, к НКТ, к другим зрительным полям), должны служить для передачи разных видов зрительной информации.
Подводя итоги, можно сказать, что при переходе от слоя к слою в поведении клеток выявляются более важные различия, чем, скажем, различия в оптимальной ориентации стимула или же в характере глазодоминантности. Наиболее заметные различия между клетками разных корковых слоев касаются сложности их реакций; это отражает тот простой анатомический факт, что одни слои расположены ближе к входу в кору, чем другие.
Колонки глазодоминантности
Группы клеток стриарной коры с различной глазодоминантностью были обнаружены раньше всего, так как они довольно крупные. Разработано много методов исследования таких группировок, поэтому они сейчас лучше всего изучены. Уже в самом начале исследований на коре обезьян стало очевидно, что всякий раз, когда микроэлектрод входит в кору перпендикулярно ее поверхности, он встречает одну за другой клетки, лучше реагирующие на стимуляцию одного и того же глаза (это показано на рис. 68). Если микроэлектрод вынуть и ввести его в другом месте в нескольких миллиметрах от предыдущего, то снова для всех встречающихся клеток доминирующим будет один глаз — тот же, что и раньше, или другой. В слое 4С, имеющем входы непосредственно от НКТ, доминирование одного глаза становится уже не относительным, а абсолютным, монопольным. В тех же слоях, которые расположены выше и ниже и, следовательно, находятся дальше в цепи синаптических переключений, более
АРХИТЕКТУРА ЗРИТЕЛЬНОЙ КОРЫ 113
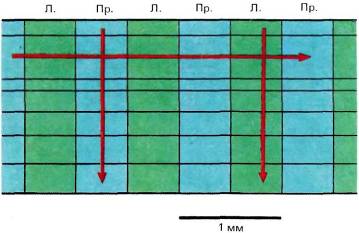
Рис. 68. Глазодоминантность остается неизменной при вертикальном продвижении микроэлектрода в стриарной коре. Если же электрод продвигается параллельно поверхности коры, то зоны с различной глазодоминантностью чередуются, причем каждая пара таких зон занимает около 1 мм.
половины всех клеток могут возбуждаться и от недоминирующего глаза. Такие клетки мы называем бинокулярными.
Если вместо того, чтобы перемещать электрод перпендикулярно поверхности коры, мы вводили его с наклоном, как можно ближе к линии, параллельной поверхности, то клетки с разной глазодоминантностью чередовались — доминирующим был то один глаз, то другой. Полный цикл этой смены соответствовал примерно одному миллиметру. Очевидно, если бы можно было видеть структуру коры сверху, она представлялась бы мозаикой, составленной из чередующихся зон с доминированием правого и левого глаза.
Чем обусловлено такое чередование, стало ясно, когда был разработан новый метод окраски, позволяющий прослеживать ветвление отдельных аксонов, приходящих из НКТ, и распределение их окончаний в коре. Ветвление одного аксона таково, что тысячи его концевых синапсов образуют в слое 4С два или три скопления шириной 0,5 мм, разделенных промежутками около 0,5 мм, как показано на рис. 69. Поскольку клетки НКТ монокулярные, каждый отдельный аксон связан в конечном счете либо с правым, либо с левым глазом. Предположим, что аксон, изображенный на рис. 69 слева, относится к волокнам от левого глаза; в этом случае каждое входящее в тот же участок коры волокно, связанное с левым глазом, будет разветвляться в тех же скоплениях окончаний величиной 0,5 мм каждое. Промежутки между этими скоплениями размером тоже 0,5 мм будут заняты концевыми разветвлениями волокон от правого глаза. Такой особый тип распределения аксонов из НКТ в слое 4С сразу позволяет объяснить строгую монокулярность клеток в этом слое.
Для того чтобы избирательно окрашивать одно и только одно волокно, требовался новый метод, и он был предложен в конце 1970-х годов. Этот метод основан на использовании аксонного транспорта — процесса, в результате которого различные материалы (белки или даже более крупные частицы) непрерывно переносятся в обоих направлениях внутри аксона. Некоторые из них транспортируются со скоростью нескольких сантиметров в час, другие — несколько миллиметров в сутки. Для того чтобы окрасить одиночный аксон, с помощью микропипетки вводят в него вещество, о котором известно, что оно переносится по аксону и окрашивает его, но не влияет на структуру клетки. В
114