

Глава 4
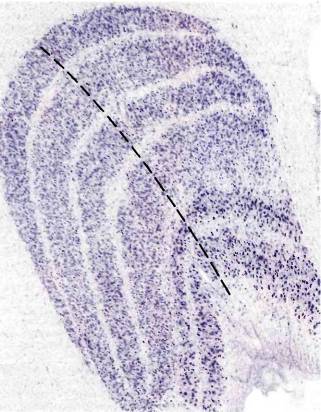
Рис. 38. Левое наружное коленчатое тело макака. Ясно видны шесть клеточных слоев. Срез сделан параллельно фронтальной плоскости; он специально окрашен для выявления тел нейронов (каждое из них выглядит как точка).
вергируют на одни и те же клетки НКТ — каждая из этих клеток получает сигналы только от какого-то одного глаза. Два множества клеток разнесены по отдельным слоям, так что в любом слое все клетки получают информацию только от одного глаза. Эти слои расположены таким образом, что проекции от правого и левого глаза чередуются. Так, в левом НКТ проекции располагаются в следующем порядке (от поверхности в глубину): левая, правая, левая, правая, правая, левая. Не совсем ясно, почему последовательность 5-го и 6-го слоев «перевернута» (иногда мне кажется, что это сделано для того, чтобы порядок проекций было труднее запомнить). У нас пока нет никакого вразумительного объяснения самого факта чередования проекций.
В целом шестислойная нейронная структура имеет одну общую топографию для всех слоев. Левые половины обеих сетчаток проецируются в слои левого НКТ (рис. 39), а правые половины — в слои правого НКТ. Любая точка
ПЕРВИЧНАЯ ЗРИТЕЛЬНАЯ КОРА 73
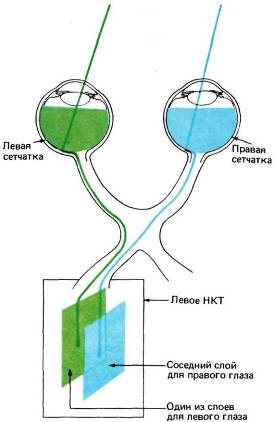
Рис. 39. При переходе от сетчатки к НКТ пространственная упорядоченность нейронов сохраняется, хотя на этом пути она временно исчезает, когда волокна собираются в пучок; в НКТ они снова «находят свои места».
в одном слое НКТ соответствует некоторой точке в поле зрения того или другого глаза. Если двигаться вдоль слоя НКТ, то соответственная точка в поле зрения будет перемещаться по некоторой траектории, определяемой характером отображения зрительного поля на НКТ. Если же двигаться перпендикулярно слоям НКТ (например, вдоль черной штриховой линии на рис. 38), как двигался бы микроэлектрод, проходя через разные слои, то соответствующие рецептивные поля клеток оставались бы в одном и том же участке поля зрения; при этом наблюдалось бы только чередование проекций от разных глаз, за исключением, конечно, того места, где идут подряд две проекции от одного глаза. Таким образом, каждая половина поля зрения шестикратно отображается на каждое из НКТ, трижды для каждого глаза, а проекции в слоях НКТ располагаются в точности друг под другом.
Наружное коленчатое тело выглядит как состоящее из двух частей. Его подразделяют на вентральные, или нижние, слои и четыре дорсальных, или верхних, слоя {вентральный — расположенный ближе к брюшной стороне тела, дорсальный — к спинной стороне). Вентральная часть НКТ образует особую структуру, так как клетки в соответствующих слоях отличаются от клеток в остальных четырех слоях — они крупнее и по-иному отвечают на зрительные стимулы. В то же время четыре слоя дорсальной части НКТ как гистологически, так и по своим электрофизиологическим свойствам сходны друг с другом. Поскольку величина клеток в этих двух отделах различна, вентральные слои
74
Глава 4
стали называть крупноклеточными, а дорсальные — мелкоклеточными.
Волокна, выходящие из шести слоев НКТ, объединяются в один широкий пучок, называемый зрительной радиацией, который идет вверх до первичной зрительной коры (см. рис. 35). Здесь эти волокна равномерно расходятся и перераспределяются так, что образуется целостная проекция с топографической организацией (это аналогично распределению волокон зрительного нерва при входе его в НКТ). И вот, наконец, мы подходим к коре.
Ответы клеток в коре
Главная тема этой главы — вопрос о том, как клетки в первичной зрительной коре отвечают на зрительные стимулы. Рецептивные поля нейронов НКТ имеют такую же организацию (разделение на центр и периферию), как и рецептивные поля ганглиозных клеток сетчатки, которые посылают свои аксоны к клеткам НКТ. Подобно ганглиозным клеткам сетчатки, нейроны НКТ различаются между собой главным образом свойствами рецептивного поля (on- или off-центр, местоположение в поле зрения) и особенностями ответов на цветовые стимулы. Возникает вопрос: а как обстоит дело с корковыми нейронами? Сходны ли они с клетками НКТ, посылающими в кору свои аксоны, или же у них появляются какие-то новые особенности? Ответ, как читатель уже должен догадаться, такой: корковые клетки действительно обладают новыми качествами, причем настолько необычными, что вплоть до 1958 года, когда их впервые стали изучать с помощью сложных световых стимулов, никто не мог даже приблизительно предсказать эти свойства.
Первичная зрительная кора (стриарная кора) представляет собой слой клеток толщиной 2 мм и площадью в несколько квадратных дюймов1. Для того чтобы дать представление о размерах этой нейронной структуры, можно привести такие цифры: если НКТ содержит полтора миллиона клеток, то стриарная кора — около 200 миллионов клеток. Анатомическая структура стриарной коры удивительно сложна, однако нет необходимости знать ее детали, чтобы понять, каким образом преобразуется здесь поступающая зрительная информация. Более подробно строение этого отдела будет рассмотрено в следующей главе, где будет обсуждаться вопрос о его функциональной архитектуре.
Как я уже говорил, процесс переработки информации в коре состоит из нескольких этапов. На первом этапе большинство клеток дает такие же ответы, как клетки НКТ. Рецептивные поля этих клеток обладают круговой симметрией. Это означает, что линия или граница (перепад освещенности) вызывает один и тот же ответ независимости от ее ориентации. Регистрировать электрическую активность корковых клеток этого уровня непросто, так как они очень малы и расположены близко друг к другу. Пока еще не ясно, отличаются ли вообще ответы этих корковых клеток от ответов клеток НКТ (точно так же как не ясно, отличаются ли реакции клеток НКТ от ответов ганглиозных клеток сетчатки). Сложность гистологического строения НКТ и коры позволяет думать, что между ними должны быть какие-то различия и что их можно будет выявить, если знать, в чем их следует искать; однако узнать это может оказаться трудным делом.
Положение еще больше усложняется, когда мы переходим к ответам клеток на следующем корковом уровне. Вероятно, эти клетки должны получать входные сигналы от нейронов предыдущего уровня, обладающих рецептивными полями с центром и периферией. Вначале нам было совсем не просто
Квадратный дюйм равен около 6,3 см2. — Прим. ред.
ПЕРВИЧНАЯ ЗРИТЕЛЬНАЯ КОРА 75
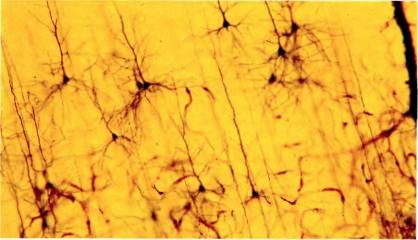
Рис. 40. Окрашенный по Гольджи срез первичной зрительной коры, на котором видно больше дюжины пирамидных клеток (но это лишь очень малая доля нейронов, содержащихся в срезе). Величина изображенного участка по вертикали — около 1 мм. (Темная полоса у правого края — кровеносный сосуд.)
выяснить, на какие зрительные стимулы отвечают эти клетки второго коркового уровня. В то время (конец 50-х годов) лишь очень немногие исследователи пытались регистрировать ответы одиночных нейронов зрительной коры. Те, кто это делал, получали противоречивые результаты. Они нашли, что клетки в зрительной коре работают, видимо, почти так же, как и в сетчатке, — были найдены как on-, так и off-клетки. Кроме того, был обнаружен еще один класс клеток, которые, казалось, вообще не отвечали на световые стимулы. Простота выявленных физиологических свойств корковых клеток на фоне просто дьявольской сложности морфологии коры ставила исследователей в тупик.
Сегодня это очень просто объяснить. Во-первых, применялись неадекватные стимулы — для активации клеток коры по привычке просто освещали всю сетчатку рассеянным светом, хотя Куффлер еще за десять лет до того показал, что такая стимуляция даже для нейронов сетчатки далеко не оптимальна. Для большинства же корковых нейронов диффузный засвет сетчатки — не только не оптимальный, но вообще полностью неэффективный стимул. Если многие клетки наружных коленчатых тел еще реагируют, хотя и слабо, на диффузный засвет, то корковые клетки, даже относящиеся к первому корковому уровню и сходные с клетками НКТ, практически не отвечают на такую стимуляцию. Таким образом, первая приходящая в голову мысль, что для активации зрительных нейронов лучше всего стимулировать все рецепторы сетчатки, оказалась совершенно неверной. Во-вторых (и это еще парадоксальнее), оказалось, что те корковые клетки, которые давали on- или off-ответы, на самом деле были не клетками, а просто аксонами клеток НКТ. Настоящие корковые нейроны в этом случае вообще не отвечали на стимуляцию! Они считали ниже своего достоинства обращать внимание на такой примитивный стимул, как диффузный свет.
Именно так обстояло дело в 1958 году, когда Торстену Визелу и мне удалось осуществить одно из первых успешных отведений электрической активности коры у кошки. В этих опытах положение кончика микроэлектрода в корковой