34 Глава 2
информации. На нашей схеме (рис. 13), разумеется, доминирует направление слева направо, от входа к выходу. Но почти во всех случаях, когда сигналы передаются от одного уровня к следующему, возвратные связи направляют их также назад, со второго уровня на первый. (Иногда мы можем догадываться, для чего такая обратная связь могла бы служить, но почти во всех случаях у нас нет на этот счет ясного понимания.) Наконец, даже в пределах какого-то одного уровня мы часто обнаруживаем богатую сеть соединений между соседними клетками. Таким образом, утверждение, что та или иная структура содержит определенное число уровней, почти всегда будет чрезмерным упрощением.
Когда я начинал работать в области неврологии в начале 50-х годов, этот основной план нервной системы был хорошо известен. Но в те дни никто не имел четкого представления о том, как понимать организацию этой информационной эстафеты по цепи уровней. Сейчас мы значительно больше знаем о способах преобразования информации в некоторых частях мозга; о других частях нам еще почти ничего не известно. Оставшиеся главы этой книги посвящены зрительной системе, которую мы сегодня понимаем лучше всего. Ниже я попытаюсь дать обзор некоторых фактов, известных нам об этой системе.
Зрительный путь
Теперь мы можем приспособить нашу первоначальную схему (рис. 13) к специальному случаю зрительного пути. Как показано на рис. 14, рецепторы и следующие два уровня находятся в сетчатке. Рецепторами служат палочки и колбочки; зрительный нерв — общий выход всей сетчатки — представляет собой пучок аксонов клеток третьего уровня, называемых ганглиозными клетками сетчатки. Между рецепторами и ганглиозными клетками имеются промежуточные нейроны, из которых наиболее важны биполярные клетки. Зрительный нерв направляется к расположенной глубоко в мозгу промежуточной станции — наружному коленчатому телу. Отсюда после всего лишь одного набора синапсов выходные сигналы направляются к стриарной коре, содержащей три или четыре уровня.
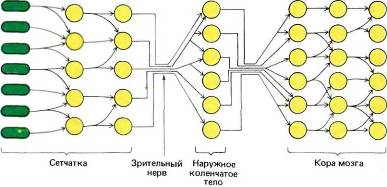
Рис. 14. Начальные уровни зрительной системы млекопитающих имеют слоистую организацию, часто встречающуюся в центральной нервной системе. Первые три уровня размещаются в сетчатке, остальные в мозгу — в наружных коленчатых телах и далее в коре мозга.
ИМПУЛЬСЫ, СИНАПСЫ И НЕЙРОННЫЕ СЕТИ 35
Рис. 15. Любой уровень в схемах на рис. 13 и 14 состоит из двумерного слоя клеток. Клетки любого уровня могут быть так плотно упакованы, что им приходится размещаться в несколько слоев; тем не менее они все принадлежат к одному уровню.
Каждую колонку приведенной схемы можно представить себе как пластину из клеток в поперечном разрезе. Если, например, вы будете смотреть налево от слоя, расположенного на странице справа, то увидите все клетки предыдущего слоя. Каждая клеточная колонка на рисунке представляет собой двумерный массив клеток, подобно тому как это показано для палочек и колбочек на рис. 15.
Начиная здесь рассуждать об отдельных уровнях, мы сразу же должны вспомнить о нашем сравнении с генеалогией. В сетчатке, как мы увидим в главе 3, минимальное число уровней от рецепторов до выхода составляет, несомненно, три, но из-за наличия двух других типов клеток некоторая часть информации проходит непрямыми путями с четырьмя или пятью уровнями от входа до выхода. Для удобства на схеме эти окольные пути, несмотря на их важность, не показаны; кроме того, упрощены также связи между нейронами. Когда я говорю о ганглиозных клетках сетчатки как об «уровне 3 или 4», это не значит, что я забыл, сколько их есть.
Для того чтобы понять характер передачи сигналов в подобного рода сети, мы можем для начала рассмотреть поведение одиночной ганглиозной клетки сетчатки. Из анатомии мы знаем, что такая клетка имеет входы от многих биполярных клеток — возможно от 12, 100 или 1000, каждая из которых тоже в свою очередь получает сигналы от сходного числа рецепторов. Как общее правило, все клетки, посылающие сигналы к какой-то одной клетке данного уровня (как, например, биполярные клетки, «питающие» одну ганглиозную клетку), тесно группируются вместе. В сетчатке нейроны, соединенные с какой-либо одной клеткой следующего уровня, обычно лежат в зоне диаметром от 1 до 2 миллиметров, и уж во всяком случае не разбросаны по всей сетчатке. Иными словами, внутри сетчатки нет соединений длиннее примерно 1— 2 миллиметров.
Если бы мы располагали подробным описанием всех связей в такой структуре и достаточными познаниями в клеточной физиологии — например, знали, какие синапсы возбуждающие, а какие тормозные, то мы в принципе могли бы вывести заключение о происходящих здесь преобразованиях информации. Однако в отношении сетчатки и коры мозга имеющиеся в нашем распоряжении сведения отнюдь не удовлетворяют этому условию. Поэтому наиболее эффективный подход к решению этой задачи заключался до сих пор в микроэлект-