

Обсуждение
Нередко высказывается мнение, что процесс органической эволюции с её общим повышением сложности биологической организации во времени противоречит второму закону термодинамики, согласно которому изменения должны были бы происходить в обратном направлении. Неоднократно утверждалось также, что второй закон термодинамики не вполне применим к органической эволюции.
Прежде всего второй закон термодинамики относится к закрытой системе, тогда как органическая эволюция происходит в системе открытой. Кроме того, организмы поглощают энергию и используют её для поддержания биологической организации клетки и всего тела. Пища обеспечивает энергию для поддержания отдельных организмов, а в конечном счете косвенно — для повышения сложности в процессе органической эволюции.
Глава 28 Направленная эволюция
Палеонтологические данные по многим группам животных и некоторым группам растений выявили у них ряд морфологических признаков, изменяющихся на протяжении длительного периода времени в каком-то определённом направлении. Такие длительные изменения в одну сторону в пределах какой-либо естественной группы представляют собой эволюционные направления, существование которых — факт, установленный палеонтологией. Об этом достаточно убедительно свидетельствуют также сравнительно-морфологические данные по многим современным группам растений, не представленным в палеонтологической летописи, но сохранившим ряд примитивных, промежуточных и продвинутых форм. Цель настоящей главы — дать достаточно подробное описание одного хорошо документированного примера эволюционного направления, а также привлечь внимание к некоторым другим примерам. Наш подход к эволюционным направлениям носит здесь описательный, а не аналитический характер; природа эволюционных направлений будет рассмотрена в гл. 31.
Примеры
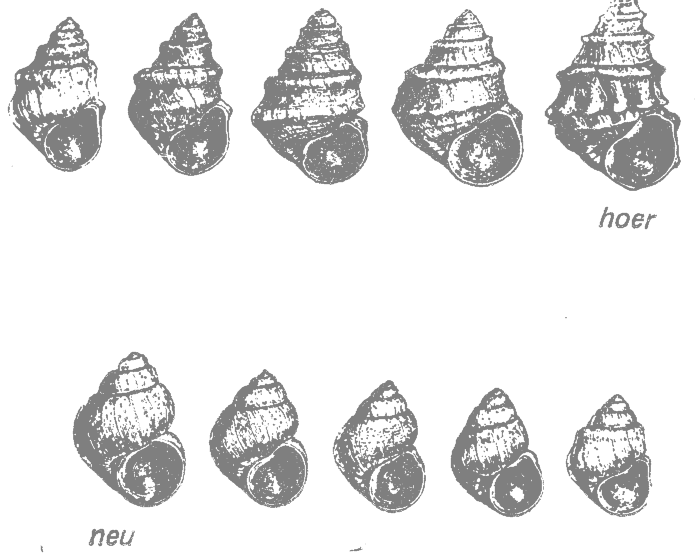
Классическим примерам эволюционного направления служит изменение закрученности и скульптурированности раковины у пресноводной улитки Paludina ( = Vivipara, Viviparidae) в юго-восточной Европе в позднем плиоцене. Серии ископаемых улиток разного возраста были найдены в озерных отложениях в Югославии и на островах Эгейского моря. Серия раковин из Югославии показана на рис. 28.1. Она начинается предковой формой P. neumayri (рис. 28.1, neu) и доходит через промежуточные формы до P. hoernesi (рис. 28.1, hoer). С течением времени закрученность раковины становится более плотной, а её скульптурированность усложняется (Abel, 1929*).
Плиоценовая P. neumayri близка к ныне живущему виду P. unicolor, который всё ещё встречается в Югославии и соседних областях. Производная форма P. hoernesi явно сильно отличается от предковой формы и её иногда выделяют в самостоятельный род Tulotoma. Её родичи больше не встречаются в Югославии, но представлены в современной фауне Северной Америки и Китая (Abel, 1929*).
Другой классический пример — увеличение рога у титанотерия (Brontotheriidae, Perissodactyla) от эоцена до олигоцена (см. гл. 33 и рис. 33.2).
Некоторые направления эволюции наблюдаются одновременно в нескольких отдельных группах, развиваясь параллельно и независимо. Так, тенденция к увеличению размеров тела наблюдается у ряда наземных млекопитающих третичного периода, например у титанотериев, лошадей и хоботных; у динозавров мезозоя; у некоторых морских животных, например у морских улиток палеозоя. Тенденция к увеличению общих размеров, так широко распространенная среди животных, была названа правилом Копа (Соре).
|
Рис. 28.1. Направления эволюции у улитки Paludina в позднем плиоцене. Эволюционный ряд тянется слева направо и от нижней серии к верхней; neu — предковая форма P. neumayri, hoer — производная форма P. hoernesi. (Из Abel, 1929 по Neumayr.*) |
Направления эволюции у цветковых растений обычно выявляют на основании сравнительно-морфологических, а не палеонтологических данных, поскольку первые в отличие от последних легко доступны (Cronquist, 1968; 1988; Тахтаджян, 1969; Stebbins, 1974; Carlquist, 1975*).
У растений, так же как и у животных, нередко можно на блюдать параллельные эволюционные направления в несвязан ных между собой группах. Обычно это изменения в сторону уменьшения размеров и упрощения морфологии, известные под общим названием редукционных рядов. Редукционные ряды одного типа, наблюдаемые во многих независимых друг от друга группах, начинаются цветками с многочисленными лепестками, тычинками и (или) плодолистиками и кончаются цветками, имеющими лишь по нескольку этих органов. Другой обычный редукционный ряд — от крупных многолетних до мелких однолетних трав. Часто такое направление у однолетних трав продолжается от перекрёстноопыляющихся форм с крупными цветками до самоопыляющихся форм с мелкими цветками.