

Распад пиримидиновых нуклеотидов
Р аспад
пиримидиновых нуклеотидов происходит
в ряде реакций:
аспад
пиримидиновых нуклеотидов происходит
в ряде реакций:
1.Отщепление 5'-фосфатной группы от ЦМФ, УМФ и ТМФ фермент 5'-нуклеотидаза.
2.Окислительное дезаминирование цитидина – аденозин-дезаминаза.
3.Дерибозилирование уридина и тимидина – нуклеозид-фосфорилаза.
4.Восстановление урацила и тимина – дегидрогеназа.
5.Гидролитическое расщепление пиримидинового кольца дигидропиримидиназой.
6.Отщепление аммиака и углекислого газа.
7.Вовлечение β-аминокислот в реакции трансаминирования,
изомеризации и далее в ЦТК
Б иосинтез пиримидиновых нуклеотидов .Регуляция биосинтеза пуриновых и пиримидиновых мононуклеотидов.
Регуляция синтеза пиримидиновых нуклеотидов
Регуляторным ферментом в синтезе пиримидиновых нуклеотидов является полифункциональный КАД-фермент. УМФ и УТФ ал-лостерически ингибируют, а ФРДФ активирует его карбамоилсинтетазную активность, тогда как активность аспартаттранскарбамоилазного домена ингибирует ЦТФ, но активирует АТФ
Этот способ регуляции позволяет предотвратить избыточный синтез не только УМФ, но и всех других пиримидиновых нуклеотидов и обеспечить сбалансированное образование всех четырёх основных пуриновых и пиримидиновых нуклеотидов, необходимых для синтеза РНК.
Регуляция синтеза пуиновых нуклеотидов
Основным показателем, от которого зависит синтез пуриновых нуклеотидов, служит концентрация ФРДФ, которая, в свою очередь, зависит от скорости его синтеза, утилизации и разрушения. Количество ФРДФ определяется доступностью рибозо-5-фосфата и активностью ФРДФ синтетазы - фермента, чувствительного к концентрации фосфата и пуриновых нуклеотидов.
Внутриклеточная концентрация ФРДФ строго регулируется и обычно низкая. ФРДФ синтета-за - аллостерический фермент. Он активируется неорганическим фосфатом (Pi) и ингибируется пуриновыми нуклеозидмоно-, ди- и трифосфатами, которые по эффективности ингибирования распределяются в следующем порядке: НМФ > НДФ > НТФ. ФРДФ служит не только субстратом, но и аллостерическим активатором второй реакции синтеза пури-нонуклеотидов de novo, которую катализирует амидофосфорибозилтрансфераза.Пуриновые нуклеотиды, особенно АМФ и ГМФ по механизму отрицательной обратной связи ингибируют амидофосфорибозилтрансфе-разу, которая катализирует первую специфическую реакцию синтеза пуриновых нуклеотидов de novo.
Биосинтез дезоксирибонуклеотидов
С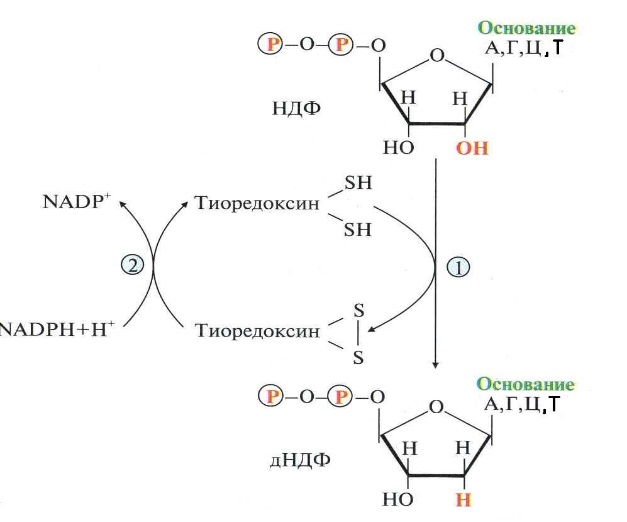 интез
дезоксирибонуклеотидов идёт с заметной
скоростью только в тех клетках, которые
вступают в S-фазу клеточного цикла и
готовятся к синтезу ДНК и делению. В
покоящихся клетках дезоксинуклеотиды
практически отсутствуют. Все
дезоксинуклеотиды, кроме тимидиловых,
образуются из рибонуклеотидов путём
прямого восстановления ОН-группы у
второго углеродного атома рибозы в
составе рибонуклеозидди-фосфатов до
дезоксирибозы.
интез
дезоксирибонуклеотидов идёт с заметной
скоростью только в тех клетках, которые
вступают в S-фазу клеточного цикла и
готовятся к синтезу ДНК и делению. В
покоящихся клетках дезоксинуклеотиды
практически отсутствуют. Все
дезоксинуклеотиды, кроме тимидиловых,
образуются из рибонуклеотидов путём
прямого восстановления ОН-группы у
второго углеродного атома рибозы в
составе рибонуклеозидди-фосфатов до
дезоксирибозы.
Реакцию восстановления НДФ в дезокси-про-изводные катализирует рибонуклеотидредуктаз-ный комплекс, в состав которого входят: собственно рибонуклеотидредуктаза (РНР), белок тиоредоксин и фермент тиоредоксинредуктаза, обеспечивающий регенерацию восстановленной формы тиоредоксина Рибонуклеотидредуктаза - олигомерный белок, состоящий из двух В1- и двух В2-субъеди-ниц, и содержит негеминовое железо в качестве кофактора.
Непосредственным донором водорода в реакции восстановления рибозы служит низкомолекулярный белок тиоредоксин. В рабочую часть этого белка входят 2 SH-группы, которые, отдавая водород, окисляются с образованием дисульфидного мостика. Второй фермент комплекса - тиоредоксинредуктаза - катализирует гидрирование окисленного тиоредоксина с использованием NADPH.