

§ 2.2 Факторы, способствующие переходу метаболизма эритроцитов из состояния с высокой концентрацией атф в устойчивое состояние с низкой концентрацией атф
Способы перевода энергетического метаболизма эритроцитов на новый уровень можно разделить на две группы: равновесные и неравновесные.
Поскольку исходное высокоэнергетическое состояние имеет определенный диапазон устойчивости по двум указанным свободным параметрам рН и nАТФ, то тактика равновесного перевода должна включать постепенное приближение (за счет регулирования процессов метаболизма) к границе устойчивости и переход за нее. Далее система сама обеспечит достижение низкоэнергетического состояния, если такое имеется. Рис. 10 иллюстрирует кинетику возможного перехода метаболизма эритроцита из одного устойчивого состояния в другое состояние в результате изменения регулирования скоростей протекания равновесных процессов.
Рис. 10. Иллюстрация кинетики перехода
метаболизма эритроцита из одного
устойчивого состояния в другое состояние
в результате изменения скоростей
протекания равновесных процессов.
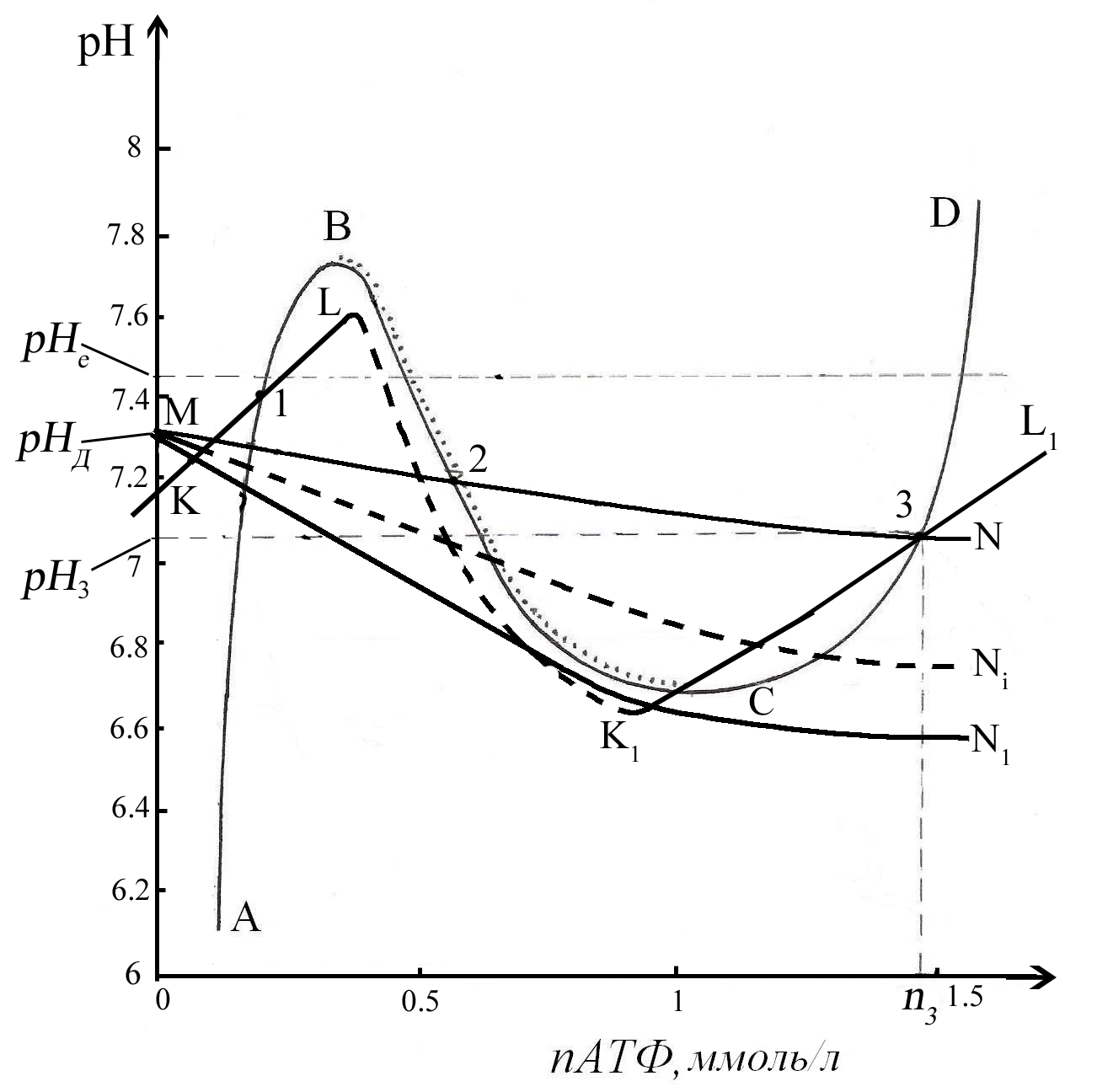
На этом рисунке на фазовой плоскости (nАТФ, рН) представлены следы кривых баланса скоростей притока и убыли основных параметров: АВСD – концентрации АТФ в цитоплазме, КLK1L1 – количества липидов С в мембране эритроцитов, МN – концентрации ионов Н+ в цитоплазме. Точка 3 на этом рисунке иллюстрирует взаимное расположение кривых, определяющее исходное состояние системы до изменения регулирования процессов. Для того, чтобы осуществился переход в область низких значений nАТФ необходимо, чтобы в результате регулирования активности равновесных процессов кривые АВСD и КLK1L1 смещались в сторону увеличения рН, а MN в сторону меньших значений этой величины. Так как для перехода важно только изменение взаимного расположения указанных кривых, то для простоты и ясности рассмотрения, можно положить, что 2 зигзагообразные кривые не меняют своего положения, а меняется только положение кривой МN. При смещении кривой MNi происходит постепенное приближение ее к границе устойчивости, что означает приближение точки Ni к точке С. Такое смещение приводит к рассогласованию рассматриваемых процессов, в результате чего появляются автоколебания параметров (рис. 11). Исследование этих колебательных процессов не является предметом рассмотрения данной работы.
Рис. 11. Рассогласование процессов
метаболизма эритроцитов, выражающееся
в колебании параметров nАТФ
и рН.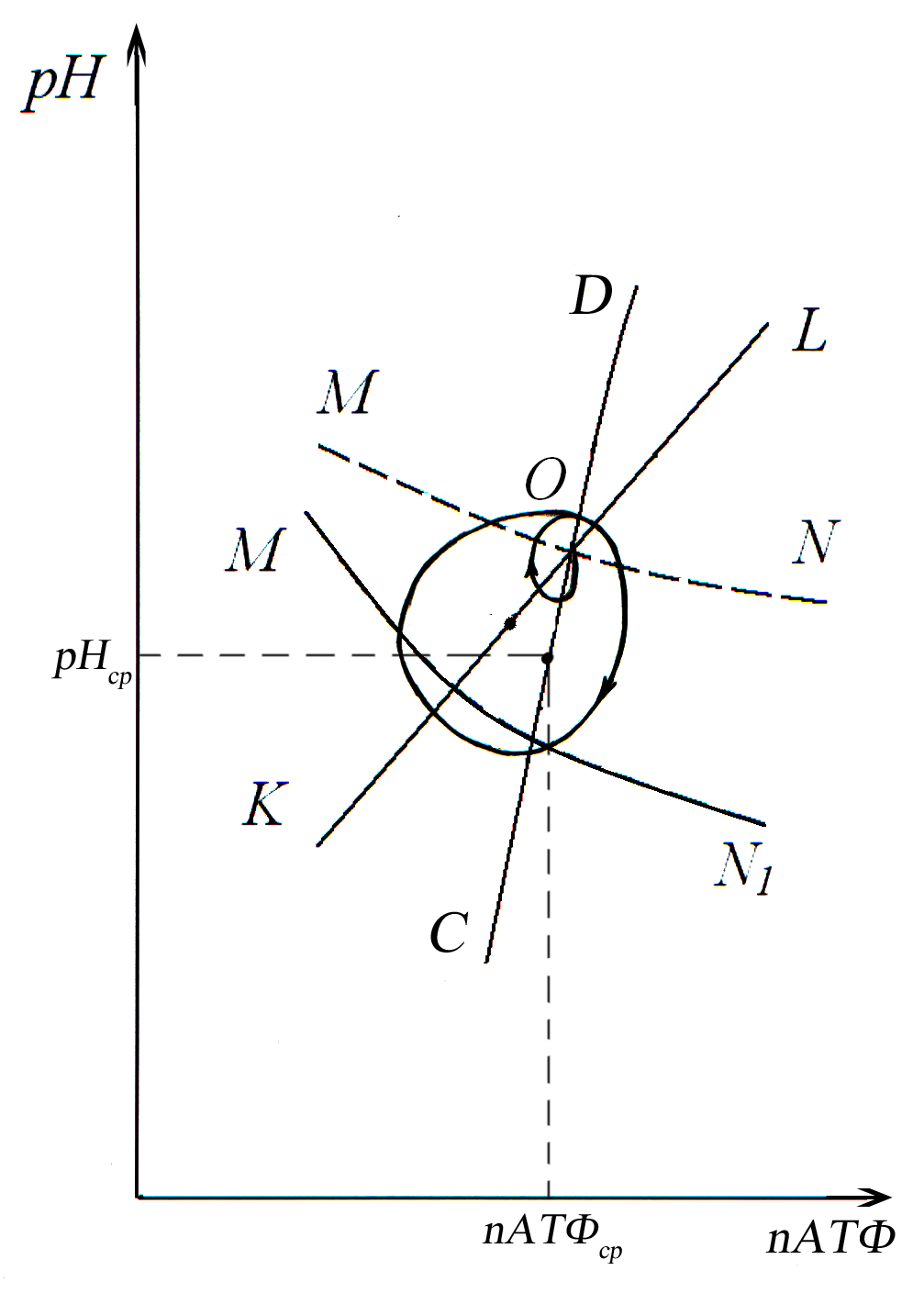
Колебания, изображенные на рис.11, облегчают переход системы в указанное выше низкоэнергетическое состояние. На этом рисунке представлены участки кривых на фазовой плоскости (nАТФ, рН), показанных на рис. 10, относящихся к исходному состоянию системы. Обозначения кривых те же, что и на предыдущем рисунке. Рассогласование процессов проявляется в том, что участки кривых не пересекаются в одной точке. За счет обратных связей между скоростями процессов и самими значениями параметров и за счет инерционности процессов, режим метаболизма системы будет характеризоваться непрерывным изменением как скоростей, так и значений самих параметров. Возникают их незатухающие колебания. Непрерывная смена значений рН и концентрации АТФ определяет на фазовой плоскости замкнутую траекторию. Процессы смены указанных величин существенно неравновесны, но среднее значение параметров за цикл «равновесно» в том смысле, что система ведет себя так, как будто строго гомеостатируется величина каждого из этих изменяющихся параметров. Соответственно, средние за цикл значения параметров рН и nАТФ для каждого отрезка кривых CD, K1L1 и MNi, находятся на каждом из них. Таким образом, достигается стабилизация процессов. Значение параметров рН, nАТФ, С и их скоростей роста V+ и убыли V- параметров стабилизированы в пределах значений точек траектории. Устойчивость будет нарушена, когда точка (nАТФср, рНср) отрезка СD, соответствующая среднему значению концентрации АТФ за цикл, достигнет точки С и перейдет за нее. Последующий переход в низкоэнергетческое состояние может сопровождаться гемолизом эритроцитов, в зависимости от взаимного расположения точек К1 и С. Чем выше ордината точки К1 по сравнению с ординатой точки С, тем выше вероятность гемолиза, так как при таком расположении может нарушится баланс скоростей уменьшения и увеличения количества в мембране липидов в сторону преобладания скоростей уменьшения количества липидов. Подробное исследование не входит в данную работу.
Рассмотрим теперь неравновесный переход энергетического метаболизма эритроцитов на новый уровень. Обратимся к рис. 12. На нем представлены те же самые кривые, что на рис. 8.
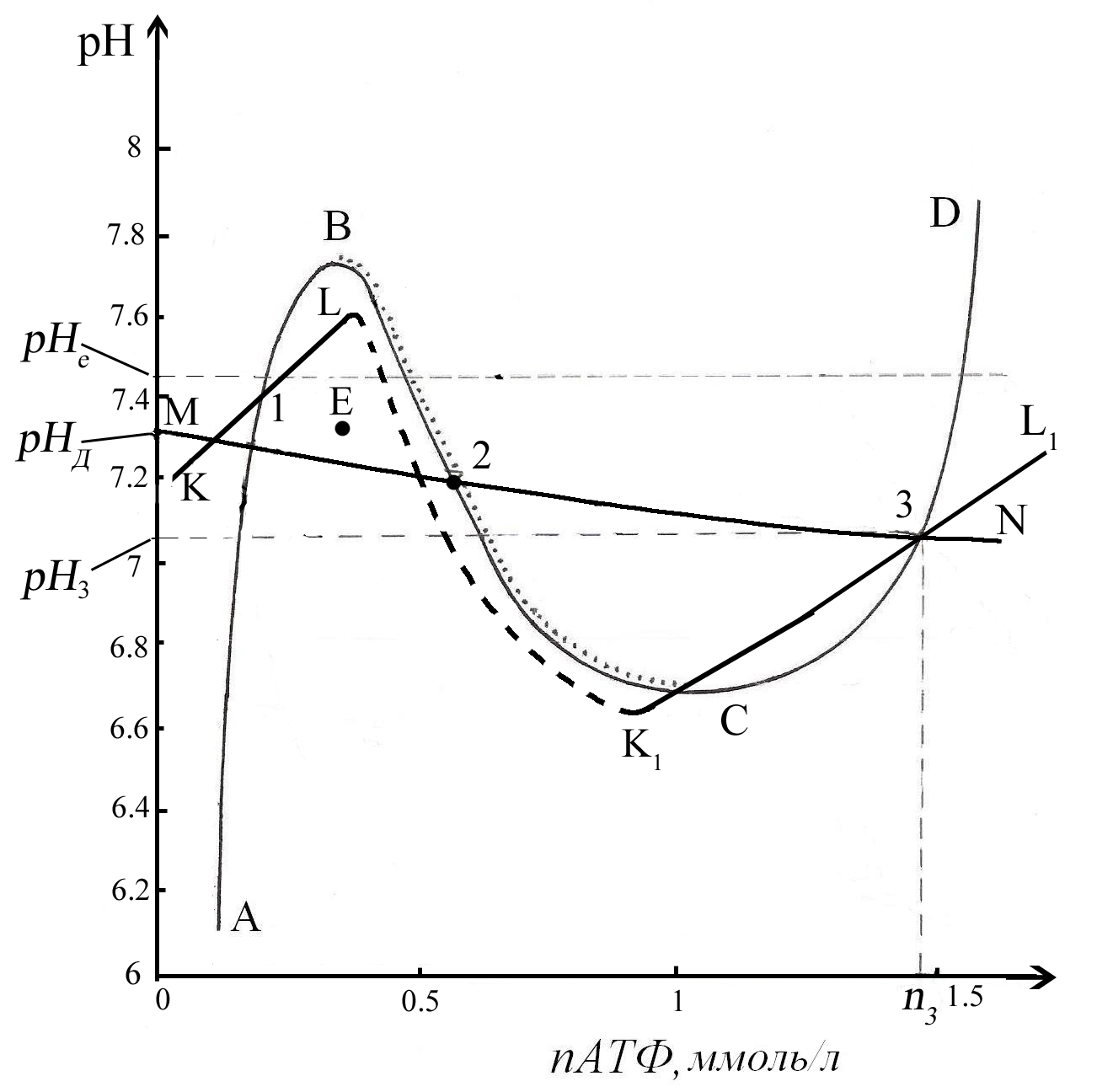
Рис. 12. Следы кривых баланса скоростей:
синтеза и расхода АТФ – кривая АВСD;
уменьшение и увеличения количества
липидов в мембране – кривая KLK1L1;
подкисления и подщелачивание цитоплазмы
– кривая МN на фазовой
плоскости (nАТФ,рН) во
всем физиологическом диапазоне
концентраций АТФ и рН. Точка Е –
предполагаемое промежуточное состояние
при неравновесных процессах, приводящих
состояние метаболизма эритроцитов к
устойчивой точке 1.
Если неравновесными процессами изменить концентрацию АТФ и рН так, что соответствующая точка будет левее отрезка ВС, то возможен переход в устойчивое состояние, соответствующее точке Е. Поскольку под кривой АВСD преобладает скорость расхода АТФ, то последующее уменьшение концентрации АТФ приближает состояние эритроцита к устойчивой точке 1. Одновременно точка оказывается под кривой KL, где доминируют процессы распада липидов. Следовательно, при движении состояния системы к устойчивой точке 1, возможно достижение критически минимального значения количества липидов, когда клетка превратится в сферу и распадется. Вероятность распада тем больше, чем дальше точка находится от кривой KL.
Итак, для проведения теоретического исследования кинетики энергетического метаболизма эритроцитов in vivo с целью выяснения наличия устойчивых состояний и их числа за основу была выбрана основная физическая модель, предложенная в работе [15]. Эта модель была нами дополнена элементами, приближающими рассмотрение метаболизма эритроцитов к in vivo. Дополненная (обобщенная) модель допускает существование второй устойчивой точки гомеостатирования параметров (рН, nАТФ и С) при низкой концентрации АТФ. Исследование выбранной модели позволило выяснить условия, при которых может происходить переход метаболизма эритроцитов из одной устойчивой точки в другую.
Для надежного установления этого факта необходимо провести экспериментальное исследование.