





































Глава 10
Мышцы и движение
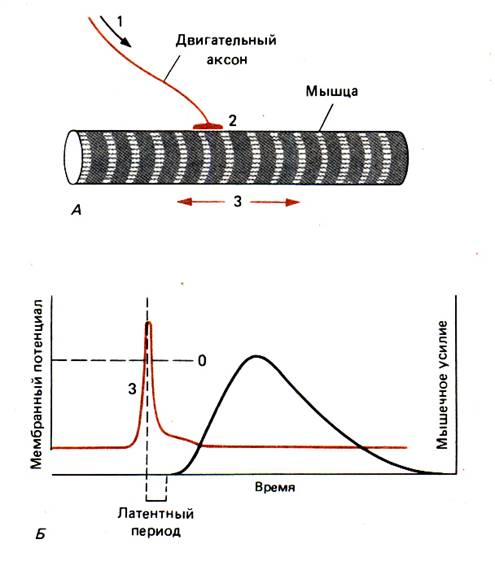
У животных есть три главных механизма генерации движения – амебоидное движение, биения ресничек и жгутиков, мышечное сокращение. Со времен античности сокращение мышц – наиболее наглядный и хорошо видимый невооруженным глазом признак жизни животного организма – особенно занимало исследователей. Гален во II веке полагал, что «животные духи», проникая по нервам в мышцу, раздувают ее, что приводит к увеличению диаметра мышцы и вместе с тем ее укорачиванию. Еще совсем недавно, в 50–х годах, утверждали, что сокращение мышцы происходит в результате укорочения линейных размеров молекул «сократительного» белка. Считалось, что такие молекулы имеют спиральную конфигурацию, изменение шага которой приводит к изменению длины. Эта гипотеза оказалась, однако, недолговечной; развитие новых методов исследования привело к полному изменению наших представлений о функционировании мышц. В итоге на сегодняшний день мы гораздо лучше разбираемся в том, как работают мышцы, чем в большинстве других областей физиологии. На основе фактов, полученных с помощью электронной микроскопии, биохимических и биофизических методов, удалось понять, как организован сократительный аппарат мышц и каким образом он обеспечивает их укорочение. Постепенно вносится ясность и в вопрос о том, каким образом электрический сигнал мембраны мышечной клетки инициирует процесс сокращения.
Как по морфологическим, так и по функциональным характеристикам мышцы разделяют на два основных типа – гладкие и поперечнополосатые. Поскольку мы наиболее осведомлены о структуре и функции поперечнополосатых мышц позвоночных животных (главным образом скелетных мышц лягушки и кролика), они и будут использованы в качестве модели при изложении большей части материала настоящей главы. Поперечнополосатые мышцы в свою очередь принято подразделять на скелетные и сердечную мышцы. Такое подразделение не основано, однако, на принципиальных различиях, поскольку у этих типов мышц сократительные механизмы почти идентичны по структуре и функции, хотя есть некоторые существенные различия в клеточной организации.
10.1. Структурные основы сокращения
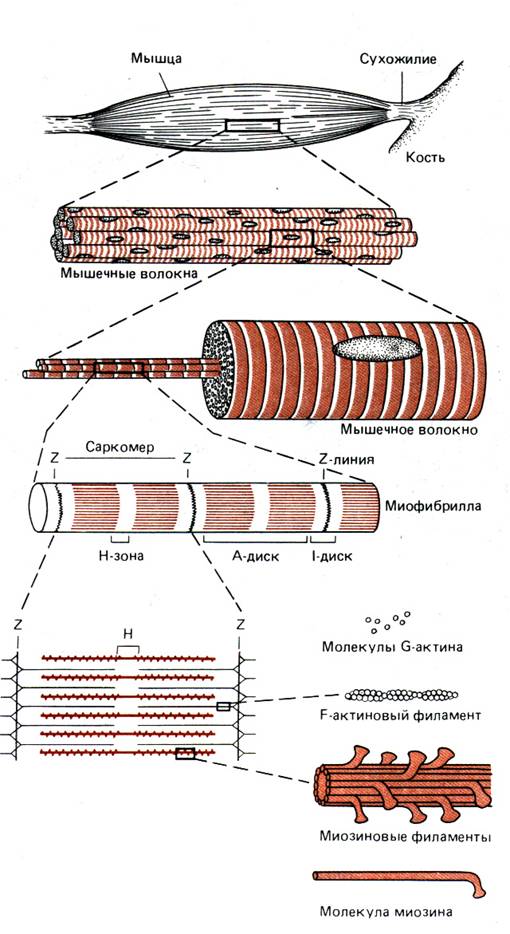
Иерархические уровни организации ткани скелетной мышцы показаны на рис. 10–1. Длинные цилиндрические многоядерные клетки, или волокна, поперечнополосатых мышц обычно начинаются от сухожилия или иной соединительной ткани, прикрепленной к одной кости, и заканчиваются в сухожилии или другой соединительной ткани, прикрепленной ко второй кости. Мышечные волокна тесно прилегают друг к другу; следовательно, они работают параллельно. Диаметр волокон поперечнополосатых мышц составляет от 5 до 100 мкм, а длина может достигать нескольких сантиметров и более. Эти необычные размеры объясняются синцитиальным происхождением мышечных волокон, которые возникают из отдельных клеток – миобластов, сливающихся в миотрубочки. Последние в свою очередь дифференцируются с образованием многоядерных, окруженных мембраной мышечных волокон. Каждое мышечное волокно состоит из множества параллельно расположенных субъединиц, называемых миофибриллами, которые включают в себя повторяющиеся в продольном направлении блоки – саркомеры, отделенные друг от друга Z–пластинками. Саркомер миофибриллы представляет собой функциональную единицу поперечнополосатой мышцы. Миофибриллы отдельного мышечного волокна связаны таким образом, что расположение саркомеров совпадает, и это создает картину «полосатости», или поперечной исчерченности, волокна при наблюдении в световой микроскоп. Тонкое строение поперечнополосатых мышц – прекрасный пример того, как структура лежит в основе функции.
|
Рис 10.1 Иерархические уровни организации скелетной мышцы (Bloom, Fawcett, 1968.)
|
|
Рис. 10.2. Саркомеры трех соседних миофибрилл на продольном тонком срезе мышцы лягушки (электронная микрофотография, × 150000). Видны I–, Н–, А–диски и Z–пластинка. Темные зерна между миофибриллами – скопления гликогена (С любезного разрешения L. D. Peachey )
|
На электронной микрофотографии (рис. 10–2) изображен продольный срез нескольких миофибрилл. Z–пластинка содержит α–актинин – один из белков, обнаруженных во всех подвижных клетках. В обоих направлениях от Z–пластинки тянутся многочисленные тонкие нити (филаменты), состоящие главным образом из белка актина. Они переплетаются с толстыми нитями белка миозина. Миозиновые филаменты образуют наиболее плотную часть саркомера – А–диск (будучи анизотропным, он сильно поляризует видимый свет). Более светлый участок в центре А–диска называют Н–зоной. В середине Н–зоны находится М–линия, в которой, как было показано, локализованы ферменты, играющие важную роль в энергетическом метаболизме (например, креатинкиназа). Участок саркомера между двумя А–дисками, называют I–диском (будучи изотропным, он почти не поляризует свет).
На поперечном срезе I–дисков видны только актиновые филаменты (рис. 10–3), а на поперечном срезе Н–зоны – только миозиновые. На участке взаимного перекрывания каждый миозиновый филамент окружен шестью актиновыми (тонкими) филаментами, которые отделяют его от других миозиновых (толстых) филаментов. В свою очередь каждый актиновый филамент окружен тремя миозиновыми.
|
Рис.10.3. организация миофибриллы. А. Схема трех саркомеров, из которых видно, как толстые и тонкие миофиламенты образуют I–, А–, Н–диски и Z–пластинки. Б. Схема расположения тонких (слева), толстых (справа) и смешанных (в центре) филаментов на разных уровнях поперечного сечения саркомера. В. Поперечный разрез наружной глазной мышцы паукообразной обезьяны (электронная микрофотография; ×100 000). Расположение саркомеров соседних миофибрилл не совпадает, что позволяет сопоставить данную микрофотографию с соответствующими схемами, изображенными выше. ( С любезного разрешения L. D.Peachey )
|
|
Рис. 10.4. Миозиновые поперечные мостики. А Электронная микрофотография, на которой поперечные мостики видны как едва различимые тяжи, направленные от миозиновых к актиновым филаментам; ×83 000.( Huxley, 1963).Б. расположение поперечных мостиков на толстом филаменте в виде двухнитчатой спирали. Показано увеличенным по отношению к микрофотографии А. ( Huxley, 1969) |
При тщательном электронно–микроскопическом исследовании на миозиновых филаментах обнаруживаются мелкие выступы, называемые поперечными мостиками (рис. 10–4, А). Во время сокращения эти выступы смыкаются с актиновыми филаментами. Поперечные мостики расположены вдоль миозиновых филаментов в виде двухнитчатой спирали (рис. 10–4, Б). Расстояние между соседними мостиками вдоль оси спирали составляет около 14,3 нм, а угол их смещения вокруг филамента 120°.
|
Рис. 10.5. Актиновые филаменты. А. Электронная микрофотография F–актиновых филаментов. Обратите внимание на расположение глобулярных мономеров в виде двухнитчатой спирали. Препарат был подготовлен для сканирующей микроскопии методом оттенения с помощью тонкой пленки металла; х 150 000. (С любезного разрешения R.B. Rice.) Б. Схематическое изображение двухнитчатой спирали F–актина, состоящей из G–актиновых мономеров. Такое строение актинового филамента предложено по результатам электронно–микроскопических исследований (например, микрофотография А) и рентгеноструктурного анализа. (Huxley, 1969.)
|