

Критерии выбора субстрата.
Критерии выбора |
Характеристики |
Производственные |
Доступность, транспортировка, стоимость, хранение |
Технологические |
Однородность, технологичность |
Биологические |
Инфицированность, селективность |
Физические |
Структура, прочность, дисперсность, влажность, влагоемкость |
Химические |
Состав, соотношение С/N, рН, питательность |
Микологические |
Рост мицелия, урожайность (биологическая эффективность) |
Экологические |
Экологическая чистота (пестициды, тяжелые металлы, радионуклиды) |
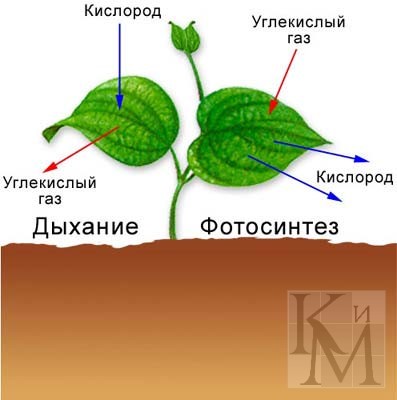
Дыхание у растений
Большинство растений в светлое время суток вырабатывают кислород, но в их клетках идёт и обратный процесс: кислород поглощается в процессе дыхания. Ночью в комнате, плотно уставленной растениями, можно наблюдать снижение концентрации кислорода и увеличение концентрации углекислого газа.
На самом деле, в живых клетках растений процесс дыхания происходит круглосуточно. Просто на свету скорость образования кислорода в результате фотосинтеза обычно превышает скорость его поглощения. Так же как и у животных, клеточное дыхание растений протекает в специальных клеточных митохондриях.
Общие принципы организации процесса дыхания на молекулярном уровне у растений и животных схожи. Однако в связи с тем, что растения ведут прикрепленный образ жизни, их метаболизм постоянно должен подстраиваться к изменяющимся внешним условиям, поэтому и их клеточное дыхание имеет некоторые особенности (дополнительные пути окисления, альтернативные ферменты).
Газообмен с внешней средой осуществляется через устьица чечевичек, трещины в коре (у деревьев).
|
Дыхание растений Дыхание растений, есть процесс более слабый, но вполне аналогичный дыханию животных; состоит в поглощении растением кислорода и выделении им углекислоты. Схема внутренних процессов может быть представлена уравнением C6H12O6+6CO2+6H2O, то есть частица сахара сжигается при помощи поглощаемого извне кислорода до углекислоты и воды. При этом процессе, выделяется теплота, что ясно заметно лишь при прорастании семян и распускании некоторых цветов. Д. растений особенно в зеленых частях маскируется гораздо более интенсивным и противоположным процессом усвоения углерода через разложение углекислоты хлорофиллом. От общего типа Д. сильно отклоняется ряд бактерий, получающих необходимую им энергию при помощи окислении минеральных веществ: серные бактерий окисляют сероводород в серную кислоту, нитрифицирующие — аммиак в азотную кислоту. |
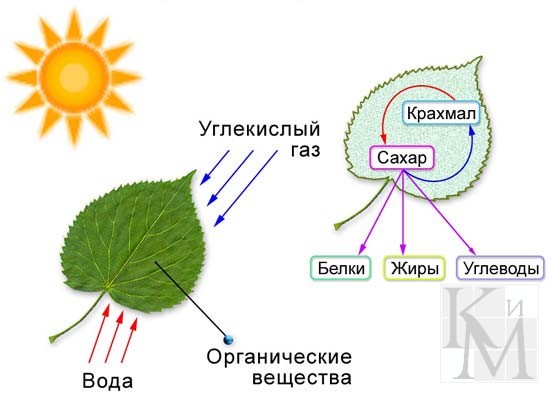
Фотосинтез
Материал из Википедии — свободной энциклопедии
Перейти к: навигация, поиск

Лист растения
Фотосинтез (от греч. φωτο- — свет и σύνθεσις — синтез, совмещение, помещение вместе) — процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов (хлорофилл у растений, бактериохлорофилл и бактериородопсин у бактерий). В современной физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндэргонических реакциях, в том числе превращения углекислого газа в органические вещества.
Типы фотосинтеза
Бесхлорофильный фотосинтез
Осуществляется археями рода Halobacterium, является наиболее примитивным типом фотосинтеза, кванты света поглощаются белком-бактериородопсином, имеющим сходство с родопсином в виде наличия ретиналя, этот тип фотосинтеза отличается отсутствием электрон-транспортной цепи, синтез АТФ осуществляется через создание электрохимического градиента протонов или ионов хлора при помощи бактериородопсиновой и галородопсиновой ионной помпы.
Хлорофильный фотосинтез
Аноксигенный
Осуществляется пурпурными и зелёными бактериями, а также геликобактериями.
Оксигенный
Оксигенный фотосинтез распространён гораздо шире. Осуществляется растениями, цианобактериями и прохлорофитами.
Этапы фотосинтеза:
фотофизический;
фотохимический;
химический.
На первом этапе происходит поглощение квантов света пигментами, их переход в возбуждённое состояние и передача энергии к другим молекулам фотосистемы. На втором этапе происходит разделение зарядов в реакционном центре, перенос электронов по фотосинтетической электронотранспортной цепи, что заканчивается синтезом АТФ и НАДФН. Первые два этапа вместе называют светозависимой стадией фотосинтеза. Третий этап происходит уже без обязательного участия света и включает в себя биохимические реакции синтеза органических веществ с использованием энергии, накопленной на светозависимой стадии. Чаще всего в качестве таких реакций рассматривается цикл Кальвина и глюкогенез, образование сахаров и крахмала из углекислого газа воздуха.
Первые процессы фотосинтеза у цианобактерий появились ещё в архейскую эру.
Пространственная локализация
Хлоропласты в клетках листа
Фотосинтез растений осуществляется в хлоропластах: обособленных двухмембранных органеллах клетки. Хлоропласты могут быть в клетках плодов, стеблей, однако основным органом фотосинтеза, анатомически приспособленным к его ведению, является лист. В листе наиболее богата хлоропластами ткань палисадной паренхимы. У некоторых суккулентов с вырожденными листьями (например, кактусы) основная фотосинтетическая активность связана со стеблем.
Свет для фотосинтеза захватывается более полно благодаря плоской форме листа, обеспечивающей большое отношение поверхности к объёму. Вода доставляется из корня по развитой сети сосудов (жилок листа). Углекислый газ поступает отчасти посредством диффузии через кутикулу и эпидермис, однако большая его часть диффундирует в лист через устьица и по листу по межклеточному пространству. Растения, осуществляющие С4 и CAM фотосинтез, сформировали особые механизмы для активной ассимиляции углекислого газа.
Внутреннее пространство хлоропласта заполнено бесцветным содержимым (стромой) и пронизано мембранами (ламеллами), которые соединяясь друг с другом образуют тилакоиды, которые в свою очередь группируются в стопки, называемые граны. Внутритилакоидное пространство отделено и не сообщается с остальной стромой, предполагается также что внутреннее пространство всех тилакоидов сообщается между собой. Световые стадии фотосинтеза приурочены к мембранам, автотрофная фиксация CO2 происходит в строме.
В хлоропластах имеются свои ДНК, РНК, рибосомы (70s типа), идёт синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь, а образуются путём деления предшествующих. Всё это позволило считать их предшественниками свободных цианобактерий, вошедших в состав эукариотической клетки в процессе симбиогенеза.
Цианобактерии, таким образом, как бы сами являются хлоропластом и в их клетке фотосинтетический аппарат не вынесен в особую органеллу. Их тилакоиды, однако, не образуют стопок, а формируют различные складчатые структуры (у единственной цианобактерии Gloeobacter violaceus тилакоиды отсутствуют вовсе, а весь фотосинтетический аппарат находится в цитоплазматической мембране, не образующей впячиваний). У них и растений также есть различия в светособирающем комплексе (см. ниже) и пигментном составе.
Световая (светозависимая) стадия
В ходе световой стадии фотосинтеза образуются высокоэнергетические продукты: АТФ, служащий в клетке источником энергии, и НАДФН, использующийся как восстановитель. В качестве побочного продукта выделяется кислород. В общем, роль световых реакций фотосинтеза заключается в том, что в световую фазу синтезируются молекула АТФ и молекулы-переносчики протонов, то есть НАДФ Н2.
Фотохимическая суть процесса
Хлорофилл имеет два уровня возбуждения (с этим связано наличие двух максимумов на спектре его поглощения): первый связан с переходом на более высокий энергетический уровень электрона системы сопряжённых двойных связей, второй — с возбуждением неспаренных электронов азота и магния порфиринового ядра. При неизменном спине электрона формируются синглетные первое и второе возбуждённое состояние, при изменённом — триплетное первое и второе.
Второе возбуждённое состояние наиболее высокоэнергетично, нестабильно и хлорофилл за 10-12 сек переходит с него на первое, с потерей 100 кДж/моль энергии только в виде теплоты. Из первого синглетного и триплетного состояний молекула может переходить в основное с выделением энергии в виде света (флуоресценция и фосфоресценция соответственно) или тепла, с переносом энергии на другую молекулу, либо, поскольку электрон на высоком энергетическом уровне слабо связан с ядром, с переносом электрона на другое соединение.
Первая возможность реализуется в светособирающих комплексах, вторая — в реакционных центрах, где переходящий в возбужденное состояние под воздействием кванта света хлорофилл становится донором электрона (восстановителем) и передаёт его на первичный акцептор. Чтобы предотвратить возвращение электрона на положительно заряженный хлорофилл, первичный акцептор передаёт его вторичному. Кроме того, время жизни полученных соединений выше чем у возбуждённой молекулы хлорофилла. Происходит стабилизация энергии и разделения зарядов. Для дальнейшей стабилизации вторичный донор электронов восстанавливает положительно заряженный хлорофилл, первичным донором же является в случае оксигенного фотосинтеза вода.
Проблемой, с которой сталкиваются при этом проводящие оксигенный фотосинтез организмы, является различие окислительно-восстановительных потенциалов воды (для полуреакции H2O → O2 (E0=+0,82 В) и НАДФ+ (E0=-0,32 В). Хлорофилл при этом должен иметь в основном состоянии потенциал больший +0,82 В чтобы окислять воду, но при этом иметь в возбуждённом состоянии потенциал меньший чем −0,32 В чтобы восстанавливать НАДФ+. Одна молекула хлорофилла не может отвечать обоим требованиям. Поэтому сформировалось две фотосистемы и для полного проведения процесса необходимо два кванта света и два хлорофилла разных типов.
Светособирающие комплексы
Хлорофилл выполняет две функции: поглощения и передачи энергии. Более 90 % всего хлорофилла хлоропластов входит в состав светособирающих комплексов (ССК), выполняющих роль антенны, передающей энергию к реакционному центру фотосистем I или II. Помимо хлорофилла в ССК имеются каротиноиды, а у некоторых водорослей и цианобактерий — фикобилины, роль которых заключается в поглощении света тех длин волн, которые хлорофилл поглощает сравнительно слабо.
Передача энергии идёт резонансным путём (механизм Фёрстера) и занимает для одной пары молекул 10−10—10−12 с, расстояние на которое осуществляется перенос составляет около 1 нм. Передача сопровождается некоторыми потерями энергии (10 % от хлорофилла a к хлорофиллу b, 60 % от каротиноидов к хлорофиллу), из-за чего возможна только от пигмента с максимумом поглощения при меньшей длине волны к пигменту с большей. Именно в таком порядке взаимно локализуются пигменты ССК, причём наиболее длинноволновые хлорофиллы находятся в реакционных центрах. Обратный переход энергии невозможен.
ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикреплённые к ним фикобилисомы — палочковидные полипептидно-пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоэритрины (с максимумом поглощения при 495—565 нм), за ними фикоцианины (550—615 нм) и аллофикоцианины (610—670 нм), последовательно передающие энергию на хлорофилл a (680—700 нм) реакционного центра.
Основные компоненты электронтранспортной цепи
Фотосистема II
Фотосистема — совокупность ССК, фотохимического реакционного центра и переносчиков электрона. Светособирающий комплекс II содержит 200 молекул хлорофилла a, 100 молекул хлорофилла b, 50 молекул каротиноидов и 2 молекулы феофитина. Реакционный центр фотосистемы II представляет собой пигмент-белковый комплекс, расположенный в тилакоидных мембранах и окружённый ССК. В нём находится димер хлорофилла a с максимумом поглощения при 680 нм (П680). На него в конечном счёте передаётся энергия кванта света из ССК, в результате чего один из электронов переходит на более высокое энергетическое состояние, связь его с ядром ослабляется и возбуждённая молекула П680 становится сильным восстановителем (E0=-0,7 В).
П680 восстанавливает феофитин, в дальнейшем электрон переносится на хиноны, входящие в состав ФС II и далее на пластохиноны, транспортируемые в восстановленной форме к b6f комплексу. Одна молекула пластохинона переносит 2 электрона и 2 протона, которые берутся из стромы.
Заполнение электронной вакансии в молекуле П680 происходит за счёт воды. В состав ФС II входит водоокисляющий комплекс, содержащий в активном центре ионы марганца в количестве 4 штук. Для образования одной молекулы кислорода требуется две молекулы воды, дающие 4 электрона. Поэтому процесс проводится в 4 такта и для его полного осуществления требуется 4 кванта света. Комплекс находится со стороны внутритилакоидного пространства и полученные 4 протона выбрасываются в него.
Таким образом, суммарный результат работы ФС II — это окисление 2 молекул воды с помощью 4 квантов света с образованием 4 протонов во внутритилакоидном пространстве и 2 восстановленных пластохинонов в мембране.
b6f или b/f-комплекс
b6f комплекс является насосом, перекачивающим протоны из стромы во внутритилакоидное пространство и создающий градиент их концентрации за счёт выделяющейся в окислительно-восстановительных реакциях электронтранспортной цепи энергии. 2 пластохинона дают перекачку 4 протонов. В дальнейшем трансмембранный протонный градиент (pH стромы около 8, внутритилакоидного пространства — 5) используется для синтеза АТФ трансмембранным ферментом АТФ-синтазой.
Фотосистема I
Светособирающий комплекс I содержит примерно 200 молекул хлорофилла.
В реакционном центре первой фотосистемы находится димер хлорофилла a с максимумом поглощения при 700 нм (П700). После возбуждения квантом света он восстанавливает первичный акцептор — хлорофилл a, тот — вторичный (витамин K1 или филлохинон), после чего электрон передаётся на ферредоксин, который и восстанавливает НАДФ с помощью фермента ферредоксин-НАДФ-редуктазы.
Белок пластоцианин, восстановленный в b6f комплексе, транспортируется к реакционному центру первой фотосистемы со стороны внутритилакоидного пространства и передаёт электрон на окисленный П700.
Циклический и псевдоциклический транспорт электрона
Помимо полного нециклического пути электрона, описанного выше, обнаружены циклический и псевдоциклический.
Суть циклического пути заключается в том, что ферредоксин вместо НАДФ восстанавливает пластохинон, который переносит его назад на b6f комплекс. В результате образуется больший протонный градиент и больше АТФ, но не возникает НАДФН.
При псевдоциклическом пути ферредоксин восстанавливает кислород, который в дальнейшем превращается в воду и может быть использован в фотосистеме II. При этом также не образуется НАДФН.
Темновая стадия
В темновой стадии с участием АТФ и НАДФН происходит восстановление CO2 до глюкозы (C6H12O6). Хотя свет не требуется для осуществления данного процесса, он участвует в его регуляции.
С3-фотосинтез, цикл Кальвина
Цикл Кальвина или восстановительный пентозофосфатный цикл состоит из трёх стадий:
карбоксилирования;
восстановления;
регенерация акцептора CO2.
На первой стадии к рибулозо-1,5-бифосфату присоединяется CO2 под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназа. Этот белок составляет основную фракцию белков хлоропласта и предположительно наиболее распространённый фермент в природе. В результате образуется промежуточное неустойчивое соединение, распадающееся на две молекулы 3-фосфоглицериновой кислоты (ФГК).
Во второй стадии ФГК в два этапа восстанавливается. Сначала она фосфорилируется АТФ под действием фосфороглицерокиназы с образованием 1,3-дифосфоглицериновой кислоты (ДФГК), затем при воздействии триозофосфатдегидрогеназы и НАДФН ацил-фосфатная группа ДФГК дефосфорилируется и восстанавливается до альдегидной и образуется глицеральдегид-3-фосфат — фосфорилированный углевод (ФГА).
В третьей стадии участвуют 5 молекул ФГА, которые через образование 4-, 5-, 6- и 7-углеродных соединений объединяются в 3 5-углеродных рибулозо-1,5-бифосфата, для чего необходимы 3АТФ.
Наконец, две ФГА необходимы для синтеза глюкозы. Для образования одной её молекулы требуется 6 оборотов цикла, 6 CO2, 12 НАДФН и 18 АТФ.
С4-фотосинтез
При низкой концентрации растворённого в строме CO2 рибулозобифосфаткарбоксилаза катализирует реакцию окисления рибулозо-1,5-бифосфата и его распад на 3-фосфоглицериновую кислоту и фосфогликолевую кислоту, которая вынужденно используется в процессе фотодыхания.
Для увеличения концентрации CO2 растения С4 типа изменили анатомию листа. Цикл Кальвина у них локализуется в клетках обкладки проводящего пучка, в клетках мезофилла же под действием ФЕП-карбоксилазы фосфоенолпируват карбоксилируется с образованием щавелеуксусной кислоты, которая превращается в малат или аспартат и транспортируется в клетки обкладки, где декарбоксилируется с образованием пирувата, возвращаемого в клетки мезофилла.
С4 фотосинтез практические не сопровождается потерями рибулозо-1,5-бифосфата из цикла Кальвина, поэтому более эффективен. Однако он требует не 18, а 30 АТФ на синтез 1 молекулы глюкозы. Это оправдывает себя в тропиках, где жаркий климат требует держать устьица закрытыми, что препятствует поступлению CO2 в лист, а также при рудеральной жизненной стратегии.
САМ фотосинтез
При CAM (англ. Crassulaceae acid metabolism — кислотный метаболизм толстянковых) фотосинтезе происходит разделение ассимиляции CO2 и цикла Кальвина не в пространстве как у С4, а во времени. Ночью в вакуолях клеток по аналогичному вышеописанному механизму при открытых устьицах накапливается малат, днём при закрытых устьицах идёт цикл Кальвина. Этот механизм позволяет максимально экономить воду, однако уступает в эффективности и С4, и С3. Он оправдан при стресстолерантной жизненной стратегии.
Значение фотосинтеза
Фотосинтез является основным источником биологической энергии, фотосинтезирующие автотрофы используют её для синтеза органических веществ из неорганических, гетеротрофы существуют за счёт энергии, запасённой автотрофами в виде химических связей, высвобождая её в процессах дыхания и брожения. Энергия, получаемая человечеством при сжигании ископаемого топлива (уголь, нефть, природный газ, торф), также является запасённой в процессе фотосинтеза.
Фотосинтез является главным входом неорганического углерода в биологический цикл. Весь свободный кислород атмосферы — биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы (кислородная катастрофа) полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни выйти на сушу.
История изучения фотосинтеза
Первые опыты по фотосинтезу были проведены Джозефом Пристли в 1770—1780-х годах, когда он обратил внимание на «порчу» воздуха в герметичном сосуде горящей свечой (воздух переставал быть способен поддерживать горение, помещённые в него животные задыхались) и «исправление» его растениями. Пристли сделал вывод что растения выделяют кислород, который необходим для дыхания и горения, однако не заметил что для этого растениям нужен свет. Это показал вскоре Ян Ингенхауз.
Позже было установлено что помимо выделения кислорода растения поглощают углекислый газ и при участии воды синтезируют на свету органическое вещество. В 1842 Роберт Майер на основании закона сохранения энергии постулировал что растения преобразуют энергию солнечного света в энергию химических связей. В 1877 В. Пфеффер назвал этот процесс фотосинтезом.
Хлорофиллы были впервые выделены в 1818 П. Ж. Пельтье и Ж. Кавенту. Разделить пигменты и изучить их по отдельности удалось М. С. Цвету с помощью созданного им метода хроматографии. Спектры поглощения хлорофилла были изучены К. А. Тимирязевым, он же, развивая положения Майера, показал что именно поглощенные позволяют повысить энергию системы, создав вместо слабых связей С-О и О-Н высокоэнергетические С-С (до этого считалось что в фотосинтезе используются жёлтые лучи, не поглощаемые пигментами листа). Сделано это было благодаря созданному им методу учёта фотосинтеза по поглощённому CO2, в ходе экспериментов по освещению растения светом разных длин волн (разного цвета) оказалось что интенсивность фотосинтеза совпадает со спектром поглощения хлорофилла.
Окислительно-восстановительную сущность фотосинтеза (как оксигенного, так и аноксигенного) постулировал Корнелис ван Ниль. Это означало что кислород в фотосинтезе образуется полностью из воды, что экспериментально подтвердил в 1941 А. П. Виноградов в опытах с изотопной меткой. В 1937 г. Роберт Хилл установил что процесс окисления воды (и выделения кислорода), а также ассимиляции CO2 можно разобщить. В 1954—1958 Д. Арнон установил механизм световых стадий фотосинтеза, а сущность процесса ассимиляции CO2 была раскрыта Мельвином Кальвином с использованием изотопов углерода в конце 1940-х, за эту работу в 1961 ему была присуждена Нобелевская премия.
В 1955 году был выделен и очищен фермент рибулозобисфосфат-карбоксилаза/оксигеназа. С4 фотосинтез был описан Ю. С. Карпиловым в 1960 и М. Д. Хэтчем и К. Р. Слэком в 1966.
Прочие факты
Морской слизень Elysia chlorotica ассимилирует хлоропласты водоросли Vaucheria litorea в клетки пищеварительного тракта. Хлоропласты способны фотосинтезировать в организме слизня в течение нескольких месяцев, что позволяет слизню жить за счет глюкозы, полученной в результате фотосинтеза. Геном слизня кодирует некоторые белки, необходимые хлоропластам для фотосинтеза.
Предполагается, что в естественной среде фотосинтезирующие бактерии могут использовать не только свет Солнца, но и другие источники света, а потому могут находиться в местах, не подвергающихся облучению звезды[2] В 2005 году Томас Битти из университета Британской Колумбии и Роберт Блейкеншип из университета Аризоны в глубоководных пробах, взятых в окрестностях глубоководного термального источника у побережья Коста Рики обнаружили серобактерию GSB1, сходную с серобактериями родов Chlorobium и Prosthecochloris, содержащую бактериохлорофилл. Они предположили, что вероятность контаминации образца невелика и, следовательно, GSB1 использует для фотосинтеза не солнечный свет (который не проникает сквозь 2,4-километровую толщу моря), а тусклый длинноволновый (~750 нм) свет, испускаемый гидротермальными источниками.
По состоянию на конец 70-х годов XX века, мощность солнечной энергии, перерабатываемой земной растительностью при фотосинтезе, всего лишь на порядок превосходила мощность всех электростанций в мире.
Корень


Ко́рень (лат. radix) — осевой, обычно подземный вегетативный орган высших растений (сосудистых растений), обладающий неограниченным ростом в длину и положительным геотропизмом. Корень осуществляет закрепление растения в почве и обеспечивает поглощение и проведение воды с растворёнными минеральными веществами к стеблю и листьям.
На корне нет листьев, в клетках корня нет хлоропластов.
Кроме основного корня, многие растения имеют боковые и придаточные корни. Совокупность всех корней растения называют корневой системой. В случае, когда главный корень незначительно выражен, а придаточные корни выражены значительно, корневая система называется мочковатой. Если главный корень выражен значительно, корневая система называется стержневой.
Некоторые растения откладывают в корне запасные питательные вещества, такие образования называют корнеплодами.
Основные функции корня
Закрепление растения в субстрате.
Всасывание, проведение воды и минеральных веществ.
Запас питательных веществ в главном корне.
Взаимодействие с корнями других растений (симбиоз), грибами, микроорганизмами, обитающими в почве (микориза, клубеньки представителей семейства Бобовые).
Вегетативное размножение.
Синтез биологически активных веществ.
У многих растений корни выполняют особые функции (воздушные корни, корни-присоски).
Происхождение корня
Тело первых вышедших на сушу растений ещё не было расчленено на побеги и корни. Оно состояло из ответвлений, одни из которых поднимались вертикально, а другие прижимались к почве и поглощали воду и питательные вещества. Несмотря на примитивное строение, эти растения были обеспечены водой и питательными веществами, так как имели небольшие размеры и жили около воды.
В ходе дальнейшей эволюции некоторые ответвления стали углубляться в почву и дали начало корням, приспособленным к более совершенному почвенному питанию. Это сопровождалось глубокой перестройкой их структуры и появлением специализированных тканей. Образование корней было крупным эволюционным достижением, благодаря которому растения смогли осваивать более сухие почвы и образовывать крупные побеги, поднятые вверх к свету. Например, у мохообразных настоящих корней нет, их вегетативное тело небольших размеров — до 30 см, обитают мхи во влажных местах. У папоротникообразных появляются настоящие корни, это приводит к увеличению размеров вегетативного тела и к расцвету этой группы в каменноугольный период.
Видоизменения и специализация корней
Корни некоторых строений имеют склонность к метаморфозу.
Видоизменения корней:
Корнеплод — утолщённый придаточный корень. В образовании корнеплода участвуют главный корень и нижняя часть стебля. Большинство корнеплодных растений двулетние. Корнеплоды состоят в основном из запасающей основной ткани (репа, морковь, петрушка).
Корневые клубни (корневые шишки) образуются в результате утолщения боковых и придаточных корней. С их помощью растение цветёт быстрее.
Корни-зацепки — своеобразные придаточные корни. При помощи этих корней растение «приклеивается» к любой опоре.
Ходульные корни — выполняют роль опоры.
Досковидные корни представляют собой боковые корни, проходящие у самой поверхности почвы или над ней, образующие треугольные вертикальные выросты, примыкающие к стволу. Характерны для крупных деревьев тропического дождевого леса.
Воздушные корни — боковые корни, растут в надземной части. Поглощают дождевую воду и кислород из воздуха. Образуются у многих тропических растений в условиях повышенной влажности.
Микориза — сожительство корней высших растений с гифами грибов. При таком взаимовыгодном сожительстве, называемом симбиозом, растение получает от гриба воду с растворёнными в ней питательными веществами, а гриб — органические вещества. Микориза характерна для корней многих высших растений, особенно древесных. Грибные гифы, оплетающие толстые одревесневшие корни деревьев и кустарников, выполняют функции корневых волосков.
Бактериальные клубеньки на корнях высших растений — сожительство высших растений с азотфиксирующими бактериями — представляют собой видоизменённые боковые корни, приспособленные к симбиозу с бактериями. Бактерии проникают через корневые волоски внутрь молодых корней и вызывают у них образование клубеньков. При таком симбиотическом сожительстве бактерии переводят азот, содержащийся в воздухе, в минеральную форму, доступную для растений. А растения, в свою очередь, предоставляют бактериям особое местообитание, в котором отсутствует конкуренция с другими видами почвенных бактерий. Бактерии также используют вещества, находящиеся в корнях высшего растения. Чаще других бактериальные клубеньки образуются на корнях растений семейства Бобовые. В связи с этой особенностью семена бобовых богаты белком, а представителей семейства широко используют в севообороте для обогащения почвы азотом.
Дыхательные корни — у тропических растений — выполняют функцию дополнительного дыхания.
Особенности строения корней
Совокупность корней одного растения называют корневой системой.
В состав корневых систем входят корни различной природы.
Различают:
главный корень,
боковые корни,
придаточные корни.
Главный корень развивается из зародышевого корешка. Боковые корни возникают на любом корне в качестве бокового ответвления. Придаточные корни образованы побегом и его частями.

Корень
Части корня
Точка роста. Участок непосредственно за кончиком корня, где клетки делятся, обеспечивая новый рост.
Зона растяжения. Участок новых клеток, образованных в точке роста и расположенных непосредственно за ней. Клетки, пока их клеточные стенки не станут жёсткими, растягиваются в длину при всасывании воды. Это растяжение толкает кончик корня дальше в почву.
Слой корневых волосков. Наиболее молодая часть эпидермиса или внешней кожицы корня. Это зона, где образуются корневые волоски. Она расположена непосредственно за зоной растяжения. Как только стенки вытянувшихся клеток становятся жёстче, самые крайние из них превращаются в слой корневых волосков. Более старая часть этого слоя (выше по корню) медленно отделяется, замещаясь слоем жёстких клеток, входящих в экзодерму (самый наружный слой коры).
Корневые волоски. Длинные выросты клеток слоя корневых волосков. Они всасывают воду и минеральные вещества.
Корневой чехлик. Слой клеток, защищающих кончик корня при росте.
Типы корневых систем
В стержневой корневой системе главный корень сильно развит и хорошо заметен среди других корней (характерна для двудольных). Разновидность стержневой корневой системы — ветвистая корневая система: состоит из нескольких боковых корней, среди которых не различают главный корень; характерна для деревьев.
В мочковатой корневой системе на ранних этапах развития главный корень, образованный зародышевым корешком, отмирает, а корневая система составляется придаточными корнями (характерна для однодольных). Стержневая корневая система проникает в почву обычно глубже, чем мочковатая, однако мочковатая корневая система лучше оплетает прилегающие частицы грунта.
Придаточные корни растут непосредственно из стебля. Они отрастают от луковицы (представляющей собой особый стебель) или от садовых черенков.
Воздушные корни. Корни, которые растут от стебля. но не проникают в землю. Они используются лазящими растениями для закрепления, как например, у плюща.
Опорные (ходульные) корни. особый тип воздушных корней. Они отрастают от стебля и затем проникают в землю, которая может быть покрыта водой. Они поддерживают тяжёлые растения, например мангры.
Зоны молодого корневого окончания
Различные части корня выполняют неодинаковые функции и различаются по внешнему виду. Эти части получили название зон.
Кончик корня снаружи всегда прикрыт корневым чехликом, защищающим нежные клетки меристемы. Чехлик состоит из живых клеток, которые постоянно обновляются. Клетки корневого чехлика выделяют слизь, она покрывает поверхность молодого корня. Благодаря слизи снижается трение о почву, её частицы легко прилипают к корневым окончаниям и корневым волоскам. В редких случаях корни лишены корневого чехлика (водные растения, некоторые растения-паразиты). Под чехликом располагается зона деления, представленная образовательной тканью — меристемой. Если эта апикальная меристема обособлена и образует только клетки корневого чехлика (как у большинства однодольных растений), её называют калиптрогеном. У большинства двудольных меристематическая ткань кончика корня сливается с меристемой, образующей зону всасывания, и называется дерматокалиптрогеном.[источник не указан 822 дня]
Клетки зоны деления тонкостенные и заполнены цитоплазмой, вакуоли отсутствуют. Зону деления можно отличить на живом корешке по желтоватой окраске, длина её около 1 мм. Вслед за зоной деления располагается зона растяжения. Она также невелика по протяжённости: составляет всего несколько миллиметров, выделяется светлой окраской и как бы прозрачна. Клетки зоны роста уже не делятся, но способны растягиваться в продольном направлении, проталкивая корневое окончание вглубь почвы. В пределах зоны роста происходит разделение клеток на ткани.
Окончание зоны роста хорошо заметно по появлению многочисленных корневых волосков. Корневые волоски располагаются в зоне всасывания, функция которой понятна из её названия. Длина её от нескольких миллиметров до нескольких сантиметров. В отличие от зоны роста участки этой зоны уже не смещаются относительно частиц почвы. Основную массу воды и питательных веществ молодые корни всасывают с помощью корневых волосков.
Корневые волоски — это выросты клеток поверхностной ткани поглощающей зоны корня растения. Они увеличивают всасывающую поверхность корня, выделяют продукты обмена; находятся чуть выше корневого чехлика. Все вместе они создают впечатление белого пушка вокруг корня. У растения, только что вынутого из почвы, всегда можно увидеть прилипшие к корневым волоскам комочки почвы. Они содержат слой протоплазмы, ядро, крупную вакуоль; их тонкие, легко проницаемые для воды оболочки плотно склеиваются с комочками почвы. Корневые волоски выделяют в почву различные вещества. Длина варьируется у разных видов растений от 0,06 до 10 мм. С увеличением влажности почвы образование замедляется; не образуются они и в очень сухой почве.
Корневые волоски появляются в виде небольших сосочков — выростов клеток. По прошествии определённого времени корневой волосок отмирает. Продолжительность его жизни не превышает 10—20 дней
Выше зоны всасывания, там, где исчезают корневые волоски, начинается зона проведения. По этой части корня вода и растворы минеральных солей, поглощённые корневыми волосками, транспортируются в вышележащие отделы растения.
Анатомическое строение корня
Для того чтобы познакомиться с системой поглощения и передвижения воды по корню, необходимо рассмотреть внутреннее строение корня. В зоне роста клетки начинают дифференцироваться на ткани и в зоне всасывания и проведения формируются проводящие ткани, обеспечивающие подъем питательных растворов в надземную часть растения.
Уже в самом начале зоны роста корня масса клеток дифференцируется на три зоны: ризодерму, кору и осевой цилиндр.
Ризодерма — покровная ткань, которой снаружи покрыты молодые корневые окончания. Она содержит корневые волоски и участвует в процессах всасывания. В зоне всасывания ризодерма пассивно или активно поглощает элементы минерального питания, затрачивая в последнем случае энергию. В связи с этим клетки ризодермы богаты митохондриями.
Экзодерма — опробковевшая покровная ткань, приходящая на смену отмирающей ризодерме.
Веламен — многослойная ризодерма, относится к первичным покровным тканям и происходит из поверхностного слоя апикальной меристемы корня. Состоит из пустотелых клеток с тонкими, опробковевшими оболочками.
Кора — образована паренхимой, обычно дифференцируется на уровне зоны растяжения. Она рыхлая и имеет систему межклетников, по которой вдоль оси корня циркулируют газы, необходимые для дыхания и поддержания обмена веществ. У болотных и водных растений межклетники коры особенно обширны. Кора является той частью корня, через которую активно проходит радиальный (ближний) транспорт воды и растворенных солей от ризодермы к осевому цилиндру. В тканях коры осуществляется активный синтез метаболитов и откладываются запасные питательные вещества.
Осевой цилиндр — представляет собой сложный комплекс из проводящей, образовательной и основной тканей.
Побег (ботаника)

Побеги подсолнечника
Побе́г (лат. córmus) — один из основных вегетативных органов высших растений, состоящий из стебля с расположенными на нём листьями и почками.
Филогенез побега
В филогенезе побеги возникли как приспособление к наземному образу жизни в результате преобразования безлистных цилиндрических органов — теломов — у риниофитов. Возникновение побегов (то есть листостебельности) — крупнейший ароморфоз в истории растительного мира на Земле. Благодаря плоской форме листьев резко возросла фотосинтезирующая поверхность, а связанное с этим увеличение транспирации способствовало развитию у растений настоящих корней как совершенных органов поглощения воды и минеральных солей.
Онтогенез побега
В онтогенезе побег развивается из почек зародыша либо из пазушной или придаточной (адвентивной) почки. Таким образом, почка является зачаточным побегом. При прорастании семени из зародышевой почечки формируется первый побег растения — его главный побег, или побег первого порядка.
Из главного побега формируются боковые побеги, или побеги второго порядка, а при повторении ветвления — третьего порядка и т. д.
Придаточные побеги формируются из придаточных почек.
Так формируется система побегов, представленная главным побегом и боковыми побегами второго и последующего порядков. Система побегов увеличивает общую площадь соприкосновения растения с воздушной средой.
В зависимости от выполняемой функции различают побеги вегетативные, вегетативно-генеративные и генеративные. Вегетативные (невидоизменённые) побеги, состоящие из стебля, листьев и почек и вегетативно-генеративные (частично видоизменённые), состоящие дополнительно из цветка или соцветия, выполняют функции воздушного питания и обеспечивают синтез органических и неорганических веществ. В генеративных (полностью видоизменённых) побегах фотосинтез чаще всего не происходит, зато там образуются спорангии, задача которых сводится к обеспечению размножения растения (к таким побегам относится и цветок).
Побег, на котором образуются цветки, называется цветоносным побегом, или цветоносом (иногда термин «цветонос» понимают в более узком смысле — как участок стебля, на котором находятся цветки).
Основные органы побега
|
Двухнедельный побег авокадо |

Для побега характерна метамерность, то есть чередование одинаковых частей — метамеров. Листья располагаются на стебле в определённом порядке, в соответствии с типом листорасположения, а участок стебля на уровне отхождения листа называется узлом. Если основание листа полностью окружает стебель, узел называется закрытым, в противном случае — открытым. Промежутки стебля между соседними узлами называют междоузлиями. Каждый повторяемый отрезок побега, который включает в себя узел с находящимся под ним междоузлием, а также расположенные на этом отрезке листья и почки называют метамером. Угол между листом и вышележащим междоузлием называют листовой пазухой, или пазухой листа.
Особенности роста побегов
В сезонном климате умеренных широт рост и развитие побегов из почек носит периодический характер. У кустарников и деревьев, а также у большинства многолетних трав это происходит один раз в году — весной или в начале лета, после чего формируются зимующие почки будущего года, а в конце лета — осенью рост побегов заканчивается. Побеги, вырастающие из почек за один вегетационный период раз в год, называют годичными. Однако формирование новых побегов из почек в течение астрономического года может быть и неоднократным. У растений, когда их вегетационный цикл состоит из нескольких периодов роста, разделённых слабо выраженными периодами покоя, нарастающие побеги называются элементарными. Так, два элементарных побега в год нередко образуются у дуба: первый — весной, второй — в середине лета; побеги летнего периода называют ивановыми побегами. У цитрусовых, чайного куста, многих тропических деревьев образуется 3—7 элементарных побегов в год. У многих многолетних трав годичные и элементарные побеги чётко не разграничены. Побеговая система травянистых поликарпиков состоит из побегов, которые после плодоношения не отмирают полностью, а их базальные участки с почками возобновления сохраняются. Это так называемые монокарпические побеги. В зависимости от длительности периода от раскрытия почки до плодоношения их делят на моноциклические (цикл развития побега завершается в течение одного вегетационного периода), дициклические (два года) и полициклические (три и более лет). Побеги, отмирающие, не доходя до цветения и плодоношения, называют побегами с неполным циклом развития.
Ветвление побегов
Поскольку побег является осевым органом, у него присутствует апикальная меристема, которая обеспечивает неограниченное нарастание в длину. Такой рост сопровождается более или менее регулярным ветвлением побега. У низших растений в результате ветвления возникает разветвлённый таллом (слоевище), у высших — образуются системы побегов и корней. Ветвление позволяет многократно увеличивать фотосинтетическую поверхность и обеспечивать растение органическими веществами.
Типы ветвления
У различных растений наблюдается несколько типов ветвления: дихотомическое, моноподиальное, симподиальное. При дихотомическом ветвлении конус нарастания расщепляется надвое. При моноподиальном ветвлении верхушечная почка функционирует в течение всей жизни, образуя главный побег (ось первого порядка), на котором в акропетальной последовательности развиваются оси второго порядка, на них — третьего порядка и т. д. При симподиальном ветвлении одна из верхних пазушных почек образует ось второго порядка, которая растёт в том же направлении, что и ось первого порядка, смещая в сторону её отмирающую часть. Впервые такое деление было предложено немецким ботаником Пфитцером в конце XIX века.
Дихотомическое ветвление

Для Псилотовидных характерно дихотомическое ветвление
Дихотомическое ветвление является наиболее примитивным типом ветвления, наблюдается как у низших растений (например у Bryophyta), так и у некоторых высших сосудистых растений (например у Lycopodiophyta, некоторых Pteridophyta,). При дихотомическом ветвлении конус нарастания делится надвое, вновь образующиеся апексы также делятся надвое и так далее.
Различают так называемое изотомическое дихотомическое ветвление (образующиеся ветви равны по размеру) и анизотомическое (при котором образующиеся ветви неравны).
Моноподиальное ветвление
Моноподиальное ветвление является следующим этапом эволюции ветвления побегов. У растений с моноподиальным типом строения побега верхушечная почка сохраняется на протяжении всей жизни побега. Моноподиальный тип ветвления часто встречается среди голосеменных растений[5], также встречается у многих покрытосеменных (например, у многих видов пальм, а также растений из семейства Орхидные — гастрохилусов, фаленопсисов и других). Некоторые из них имеют единственный вегетативный побег (например, Фаленопсис приятный).
Моноподиальные растения — термин, наиболее часто используемый в описании растений тропической и субтропической флоры, а также в научно-популярной литературе по комнатному и оранжерейному цветоводству.
Моноподиальные растения могут существенно различаться внешне. Среди них есть розеточные, с удлиненным побегом, кустовидные.

Phalaenopsis braceana. Ботаническая иллюстрация из книги J. D. Hooker; «A Century of Indian Orchids» 1895 г.

Phalaenopsis amabilis. Ботаническая иллюстрация из книги Frederick Sander, «Reichenbachia I» 1888 г.

Gastrochilus pseudodistichus. Ботаническая иллюстрация из книги G. King and R. Pantling. «The Orchids of the Sikkim-Himalaya» 1898 г.

Vanda cristata. Ботаническая иллюстрация из книги Curtis's botanical magazine vol. 73 ser. 3 nr. 3 tab. 4304, 1847 г.
Симподиальное ветвление
У растений с симподиальным типом строения побега верхушечная почка, закончив развитие, отмирает или дает начало генеративному побегу. После цветения этот побег больше не растет, а у его основания начинает развиваться новый[6]. Структура побега у растений с симподиальным типом ветвления сложнее, чем у растений с моноподиальным типом; симподиальное ветвление является эволюционно более продвинутым типом ветвления.[источник не указан 857 дней] Слово «симподиальный» образовано от др.-греч. συν- («вместе») и πούς («нога»).
Симподиальное ветвление характерно для многих покрытосеменных растений: например, для лип, ив[7] и многих орхидных.
У орхидных, помимо верхушечных, у части симподиальных орхидей образуются и боковые соцветия, развиваясь из почек, расположенных у основания побега (Пафиния гребенчатая). Часть побега, прижатая к субстрату, называется ризомой. Располагается, как правило, горизонтально и не имеет настоящих листьев, только чешуевидные. Редуцированной, почти неразличимой ризома бывает у многих масдеваллий, дендробиумов и онцидиумов; хорошо различимой и утолщённой — у каттлей и лелий, удлинённой — у бульбофиллюмов и целогин, достигая 10 и более сантиметров. Вертикальная часть побега часто бывает утолщённой, образующей так называемый туберидий, или псевдобульбу. Псевдобульбы могут быть различной формы — от почти сферической до цилиндрической, конусовидной, булавовидной и удлинённой, напоминающей стебли тростника. Псевдобульбы являются запасающими органами.
Симподиальные растения — термин, наиболее часто используемый в описании растений тропической и субтропической флоры, а также в научно-популярной литературе по комнатному и оранжерейному цветоводству.

Bulbophyllum grandiflorum. Curtis's botanical magazine vol. 127 ser. 3 nr. 57 tab. 7787, 1901 г.

Cirrhopetalum macraei. Curtis's botanical magazine vol. 127 ser. 3 nr. 57 tab. 7787, 1901 г.

Oncidium dasystyle. Curtis's botanical magazine vol. 127 ser. 3 nr. 57 tab. 7787, 1901 г.

Dendrobium senile. Curtis's botanical magazine vol. 127 ser. 3 nr. 57 tab. 7787, 1901 г.
На первых этапах эволюции верхушечные меристемы обеих ветвей каждого развилка растут с одной и той же скоростью, что приводит к образованию одинаковых или почти одинаковых дочерних ветвей. Подобная равная дихотомия (изотомия) представляет собой первоначальный тип дихотомического ветвления. Она была характерна для некоторых риниофитов, но встречается и у некоторых современных плауновидных и папоротниковидных, а также у псилота (Psilotum). В результате же неравного роста двух дочерних ветвей, когда одна из ветвей несколько опережает другую, равная дихотомия переходит в неравную дихотомию (анизотомию), очень хорошо выраженную у примитивного вымершего девонского рода хорнеофит (Horneophyton).
Эволюция типов ветвления
Видоизменения побегов (метаморфоз)
Побег — самый изменчивый по внешнему облику орган растения. Это связано не только с общей мультифункциональностью вегетативных органов, возникшей в процессе эволюции, но и с изменениями, происходящими в процессе онтогенеза растений, обусловленными адаптацией к разнообразию условий окружающей среды, а у культурных растений — под воздействием человека.
Диапазон метаморфозов побега очень широк: от небольшого уклонения от типичного строения до сильно изменённых форм. Метаморфизироваться могут главные и боковые побеги растений, почки и листья.
Основной тип побега зелёного растения — надземный (воздушный) ассимилирующий побег, несущий на оси зелёные листья срединной формации. Однако и ассимилирующие побеги не одинаковы. Нередко наряду с основной функцией фотосинтеза у этих побегов выступают и другие: отложение запасов и опорная функция (большей частью в многолетних стеблях), вегетативное размножение (ползучие побеги, плети).
Видоизменение подземных побегов
Побеги, живущие под землёй, под влиянием комплекса условий, резко отличных от наземной среды, почти полностью утратили функции фотосинтеза и приобрели другие не менее важные жизненные функции, такие как способствование перенесению неблагоприятного периода, запасания питательных веществ, вегетативного возобновления и размножения растений. К подземным видоизменённым побегам относятся: корневище, каудекс, подземные столон и клубень, луковица, клубнелуковица.
Корневище, или ризом — подземный побег с чешуевидными листьями низовой формации, почками и придаточными корнями. Толстые, сильно разветвлённые ползучие корневища характерны для пырея, короткие и довольно мясистые — для купены, ириса, очень толстые — для кубышки, кувшинки.
Каудекс — многолетний орган побегового происхождения многолетних трав и полукустарничков с хорошо развитым стержневым корнем, сохраняющимся в течение всей жизни растения. Вместе с корнем он служит местом отложения запасных веществ и несёт на себе множество почек возобновления, часть которых могут быть спящими. Каудексовых растений много среди зонтичных (бедренец, ферула), бобовых (люцерны, люпины), сложноцветных (одуванчик, полыни, василёк шероховатый).
Подземный столон — однолетний удлинённый тонкий подземный побег с недоразвитыми чешуевидными листьями. На утолщённых концах столонов растения могут накапливать запасные вещества, образуя клубни или луковицы (картофель, седмичник, адокса).
Стеблевой клубень — видоизменённый побег с ярко выраженной запасающей функцией стебля, наличием чешуевидных листьев, которые быстро сшелушиваются, и почек, формирующихся в пазухах листьев и называемых глазками (картофель, топинамбур).
Луковица — подземный (реже надземный) сильно укороченный специализированный побег, в котором запасные вещества откладываются в чешуях листовой природы, а стебель преобразован в донце. Луковица — типичный орган вегетативного возобновления и размножения. Луковицы свойственны однодольным растениям из семейства Лилейные (лилия, тюльпан, лук), Амариллисовые (амариллис, нарцисс, гиацинт) и др. Как исключение, они встречаются и у двудольных — у некоторых видов кислицы и жирянки.
|
Ixia sp.: начинающая прорастать клубнелуковица |

Видоизменения надземных побегов
Необычный образ жизни и/или приспособления к особым условиям существования растений приводят к различным видоизменениям побегов. При этом побеги могут служить не только для хранения питательных веществ, воспроизведения и размножения растений, но и выполнять другие функции. Нередки случаи, когда видоизменяется не весь побег, а только его листья, причём некоторые их метаморфозы внешне и функционально схожи с метаморфозами побега (колючки, усики).
Колючка — сильно одревесневающий безлистный укороченный побег с острой верхушкой. Колючки побегового происхождения выполняют главным образом защитную функцию. У дикой яблони, дикой груши, крушины слабительной (Rhamnus cathartica) в колючки превращаются укороченные побеги, имеющие ограниченный рост и оканчивающиеся остриём. У гледичии (Gleditschia triacanthos) мощные разветвлённые колючки образуются на стволах из спящих почек. Многие виды боярышника имеют колючки, формирующиеся из пазушных почек листьев, что топографически соответствует боковым побегам.
Усик — жгутовидный ветвистый или неразветвлённый побег метамерного строения, в типичном случае лишённый листьев. Стеблевые усики, как узкоспециализированный побег, выполняют опорную функцию. Неветвящаяся прямая часть усика представляет собой первое междоузлие пазушного побега, а закручивающаяся часть соответствует листу. Усики характерны для растений, которые не могут самостоятельно поддерживать вертикальное (ортотропное) положение. У страстоцвета голубого (Passiflora edulis) усики — это пазушные побеги моноподия, находящиеся в пазухах листьев, как и цветки. Ветвистые усики винограда (Vitis) представляют собой концы боковых побегов монохазия. У девичьего винограда (Parthenocissus) концы усиков преобразованы в прикрепительные присоски. Усики побегового происхождения образовываются также у представителей семейства Тыквенные (огурец, дыня, арбуз, тыква).
Кладодий — видоизменённый боковой побег, обладающий способностью к длительному росту, с зелёными плоскими длинными стеблями, выполняющими функции листа. Как орган фотосинтеза, кладодий имеет хорошо развитую хлорофиллоносную ткань, расположенную под эпидермой. К растениям с кладодиями относятся мюленбекия плоскоцветочная (Muhlenbekia platyclada), кактус-декабрист (Zygocactus truncates), кармихелия южная (Carmichaelia australis), коллеция (Colletia cruciata) и опунция (Opuntia).
Филлокладий — видоизменённый листовидно уплощённый боковой побег, обладающий ограниченным ростом и выполняющим функции листа. Филлокладии развиваются из боковых почек, поэтому всегда находятся в пазухе небольшого плёнчатого или чешуевидного листа. Выполняя функцию фотосинтеза, побеги филлокладиев и внешне приобретают сходство с листом, которое проявляется в ограниченном росте и полной потере метамерной структуры. Явление филлокладии свойственно таким растениям, как иглица, смела, видам родов спаржа (Asparagus), филлантус (Phyllanhtus). Филлокладии встречаются не только у покрытосеменных, но и у некоторых голосеменных, в частности, у хвойного растения из семейства Ногоплодниковые — филлокладуса.
Розеточные побеги — ненормальные побеги, образующиеся на соснах вследствие вреда, нанесённого соснам некоторыми вредными насекомыми, например, бабочкой-монашенкой и др.; такие побеги чрезвычайно коротки и имеют пучки коротких и широких игл.
Стебель


Побеги спаржи, как и «колонны» кактуса Carnegiea gigantea суть стебли
Сте́бель — удлинённый побег высших растений, служащий механической осью, также выполняет роль производящей и опорной базы для листьев, почек, цветков.
Классификация стеблей
Виды стеблей по расположению относительно уровня почвы:
надземные
подземные
Виды стеблей по степени одревеснения
травянистые
деревянистые (например, ствол — главный многолетний стебель дерева; стебли кустарников называют стволиками)
Виды стеблей по направлению и характеру роста:
прямостоячие (например, подсолнечник)
лежачие (стелющиеся) — стебли лежат на поверхности почвы, не укореняясь (вербейник монетчатый)
приподнимающиеся (восходящие) — нижняя часть стебля лежит на поверхности почвы, а верхняя поднимается вертикально (сабельник)
ползучие — стебли стелются по земле и укореняются благодаря образованию в узлах придаточных корней (будра плющевидная)
цепляющиеся (лазящие) — прикрепляются к опоре с помощью усиков (горох)
вьющиеся — тонкие стебли, обвивающие опору (луносемянник)
Виды стеблей по форме поперечного сечения:
округлые
сплюснутые
трёх—, четырёх—, многогранные (гранистые)
ребристые
бороздчатые (желобчатые)
крылатые — стебли, у которых по острым граням тянутся плоские травянистые выросты (чина лесная) или низбегающие на стебель основания листьев (окопник лекарственный).
Видоизменения стеблей
Надземные видоизмененные побеги:
почки
колючки
усики
туберидии (псевдобульбы)
Почки защищают молодой побег от холода, жары и других неблагоприятных условий среды. Почка — это побег в зачаточном состоянии. В благоприятных условиях из неё развивается побег.
Почки бывают:
верхушечные — они обеспечивают рост стебля в длину
пазушные — из них образуются боковые побеги
цветочные — из них развиваются цветки
придаточные — из них развиваются придаточные побеги — поросль. Придаточные побеги берут начало от спящих почек
Подземные видоизменённые побеги:
корневища
клубни
луковицы
стеблекорневой тубероид
Стеблекорневой тубероид — запасающий, ежегодно заменяющийся подземный орган, сочетающий в своем анатомическом строении корневые и стеблевые структуры. Характерен для представителей Orchidoidaceae. Строение стеблекорневого тубероида имеет важное таксономическое значение на уровне родов и видов.
Корневище не имеет корневого чехлика и корневых волосков. На нём есть листья в виде чешуйчатой плёнки, в пазухе размещаются почки. Из одной части этих почек образуется надземный побег, а из другой — подземный. Из верхушечной почки корневища непрерывно образуется подземный корневищный стебель, а из боковых почек — надземные стебли. Корневища отличаются высокой жизнеспособностью. Некоторые растения размножают корневищами, для этого корневище делят на части с почками, закапывают в землю.
Клубни размещаются на верхушке столонов, образуемых у основания подземных стеблей растения. Органические вещества в них накапливаются в виде крахмала. Эта часть стебля утолщается и превращается в клубень. На поверхности клубня картофеля имеется много углублений, называемых глазками. Они расположены по спирали. В каждом глазке по 2—3 почки. Картофель размножают клубнями или разделёнными частями клубня с почками.
Луковица — укороченный подземный побег. Луковицы бывают грушевидной, яйцевидной и шаровидной формы. Донце луковицы — это укороченный стебель. На донце расположены видоизменённые листья — чешуйки. Внутренние чешуйки богаты сахаристыми веществами. На нижней части донца образуется мочковатая корневая система. В пазухе мясистых чешуек, прикреплённых к донцу, очень часто есть почки, из которых развиваются новые луковички — детки.
Ветвление стебля
Это процесс образования побегов нового годичного прироста из почек на побегах прошлого года.
Лист

Лист

Осенние листья
Ли́ст (множ. ли́стья, собир. листва́) — в ботанике наружный орган растения, основной функцией которого является фотосинтез. Для этой цели лист, как правило, имеет пластинчатую структуру, чтобы дать клеткам, содержащим специализированный пигмент хлорофилл в хлоропластах, получить доступ к солнечному свету. Лист также является органом дыхания, испарения и гуттации (выделения капель воды) растения. Листья могут задерживать в себе воду и питательные вещества, а у некоторых растений выполняют и другие функции.
Анатомия листьев
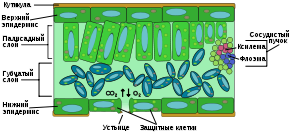
Строение листовой пластинки. Показаны палисадная (сверху, плотно упакованные клетки) и губчатая (снизу, рыхло расположенные клетки) части мезофилла, расположенные между верхним и нижним эпидермальными слоями
Как правило, лист состоит из следующих тканей:
Эпиде́рмис — слой клеток, которые защищают от вредного воздействия среды и излишнего испарения воды. Часто поверх эпидермиса лист покрыт защитным слоем восковидного происхождения (кутикулой).
Мезофи́лл, или паренхи́ма — внутренняя хлорофиллоносная ткань, выполняющая основную функцию — фотосинтез.
Сеть жи́лок, образованных проводящими пучками, состоящими из сосудов и ситовидных трубок, для перемещения воды, растворённых солей, сахаров и механических элементов.
У́стьица — специальные комплексы клеток, расположенные, в основном, на нижней поверхности листьев; через них происходит испарение воды и газообмен.
Эпидерма
Эпидерма — наружный слой многослойной структуры клеток, покрывающий лист со всех сторон; пограничная область между листом и окружающей средой. Эпидерма выполняет несколько важных функций: защищает лист от излишнего испарения, регулирует газообмен с окружающей средой, выделяет вещества обмена и в некоторых случаях впитывает воду. Большинство листьев имеют дорсовентральную анатомию: верхняя и нижняя поверхности листа имеют различную структуру и выполняют разные функции.
Эпидерма обычно прозрачна (в её строении отсутствуют либо присутствуют в недостаточном количестве хлоропласты) и снаружи покрыта защитным слоем восковидного происхождения (кутикула), который препятствует испарению. Кутикула нижней части листа, как правило, тоньше, чем на верхней, и толще в биотопах с засушливым климатом по сравнению с теми биотопами, где недостаток влаги не ощущается.
В состав ткани эпидермы входят следующие типы клеток: эпидермальные (или двигательные) клетки, защитные клетки, вспомогательные клетки и трихомы. Эпидермальные клетки самые многочисленные, крупные и наименее приспособленные. У однодольных растений они более растянуты, чем у двудольных. Эпидерма покрыта порами, называемыми устьицами, которые являются частью целого комплекса, состоящего из поры, со всех сторон окружённой содержащими хлоропласт защитными клетками, и от двух до четырёх побочных клеток, в которых хлоропласт отсутствует. Этот комплекс регулирует испарение и газообмен листа с окружающей средой. Как правило, количество устьиц на нижней части листа больше, чем на верхней. У многих видов поверх эпидермиса вырастают трихомы.
Мезофилл
Бо́льшую часть внутренности листа между верхним и нижним слоями эпидермиса составляет паренхима (основная ткань), или мезофилл. В норме мезофилл образован хлорофиллсинтезирующими клетками, поэтому употребляется и синонимичное название — хлоренхима. Продукт фотосинтеза называется фотосинтат.
У папоротников и большинства цветковых растений мезофилл разделён на два слоя:
Верхний, палисадный слой плотно упакованных, вертикально-расположенных клеток прямо под верхним слоем эпидермиса; толщиной в одну или две клетки. Клетки этого слоя содержат гораздо больше хлоропластов, чем в нижележащем губчатом слое. Длинные клетки цилиндрической формы, как правило, уложены в один — пять слоёв. Они, находясь близко к границе листа, расположены оптимально для получения солнечного света. Небольшие промежутки между клетками используются для поглощения углекислого газа. Промежутки должны быть достаточно малыми, чтобы поддерживать капиллярное действие по передаче воды. Растения должны адаптировать свою структуру для оптимального получения света при различных природных состояниях, таких как солнце или тень — солнечные листья имеют многослойный палисадный слой, в то время как теневые и старые, лежащие близко к земле листья имеют только один слой.
Клетки нижнего, губчатого слоя упакованы рыхло и, вследствие этого, губчатая ткань обладает большой внутренней поверхностью благодаря развитой системе межклетников, сообщающихся друг с другом и с устьицами. Рыхлость губчатой ткани играет важную роль в газообмене листа кислородом, углекислым газом и парами воды.
Листья обычно окрашены в зелёный цвет благодаря хлорофиллу — фотосинтезирующему пигменту, находящемуся в хлоропластах — зелёных пластидах. Растения, у которых ощущается недостаток либо отсутствие хлорофилла, не могут фотосинтезировать.
В некоторых случаях (см. Растительные химеры) в результате соматических мутаций возможно образование участков мезофилла мутантными клетками, не синтезирующими хлорофилл, при этом листья таких растений имеют пёструю окраску, обусловленную чередованием участков нормального и мутантного мезофилла (см. Пестролистность).
Растения в умеренных и северных широтах, а также в сезонно-сухих климатических зонах могут быть листопадными, то есть их листья с приходом неблагоприятного сезона опадают либо отмирают. Этот механизм имеет название сбрасывания или опадания. На месте опавшего листа на веточке образуется рубец — листовой след. В осенний период листья могут окраситься в жёлтый, оранжевый или красный цвет, так как с уменьшением солнечного света растение уменьшает выработку зелёного хлорофилла, и лист приобретает окраску вспомогательных пигментов, таких как каротиноиды и антоцианы.
Жилки
Жилки листа являются сосудистой тканью и расположены в губчатом слое мезофилла. По рисунку разветвления жилки, как правило, повторяют структуру разветвления растения. Жилки состоят из ксилемы — ткани, служащей для проведения воды и растворённых в ней минеральных веществ, и флоэмы — ткани, служащей для проведения органических веществ, синтезируемых листьями. Обычно ксилема лежит поверх флоэмы. Вместе они образуют основную ткань, называемую сердцевиной листа.
Морфология листа

Хвоя ели канадской (Picea glauca)
Лист покрытосеменных растений состоит из черешка (стебелька листа), листовой пластинки (лопасти) и прилистников (парных придатков, расположенных по обеим сторонам основания черешка). Место, где черешок примыкает к стеблю, называется влагалищем листа. Угол, образованный листом (черешком листа) и вышерасположенным междоузлием стебля, называется пазухой листа. В пазухе листа может образоваться почка (которая в этом случае называется пазушной почкой), цветок (называется пазушным цветком), соцветие (называется пазушным соцветием).
Не все растения имеют все вышеперечисленные части листьев, у некоторых видов парные прилистники чётко не выражены либо отсутствуют; может отсутствовать черешок, а структура листа может не быть пластинчатой. Огромное разнообразие строения и расположения листьев перечислены ниже.
Внешние характеристики листа, такие как форма, края, волосистость и т. д., очень важны для идентификации вида растения, и ботаники создали богатую терминологию для описания этих характеристик. В отличие от других органов растения, листья являются определяющим фактором, так как они вырастают, образуют определённый рисунок и форму, а потом опадают, в то время как стебли и корни продолжают свой рост и видоизменение в течение всей жизни растения и по этой причине не являются определяющим фактором.
Примеры терминологии, используемой в классификации листьев, можно найти в иллюстрированной английской версии Викиучебника.
Основные типы листьев
Листовидный отросток у определённых видов растений, таких как папоротники.
Листья хвойных деревьев, имеющих игловидную либо шиловидную форму (хвоя).
Листья покрытосеменных (цветковых) растений: стандартная форма включает в себя прилистник, черешок и листовую пластинку.
Плауновидные (Lycopodiophyta) имеют микрофилловые листья.
Обвёрточные листья (тип, встречающийся у большинства трав)
Расположение на стебле
По мере роста стебля листья располагаются на нём в определённом порядке, который обусловливает оптимальный доступ к свету. Листья появляются на стебле по спирали, как по часовой стрелке, так и против неё, под определённым углом расхождения. В угле расхождения замечена точная последовательность Фибоначчи: 1/2, 2/3, 3/5, 5/8, 8/13, 13/21, 21/34, 34/55, 55/89. Такая последовательность ограничена полным оборотом в 360°, 360° x 34/89 = 137,52 или 137° 30' — угол, в математике известный под названием золотой угол. В последовательности номер даёт количество оборотов до того момента, пока лист не вернётся в своё первоначальное положение. Нижеприведённый пример показывает углы, при которых листья расположены на стебле:
Очередные листы расположены под углом 180° (или 1/2)
120° (или 1/3): три листа в обороте
144° (или 2/5): пять листьев за два оборота
135° (или 3/8): восемь листьев за три оборота
Обычно же листорасположение описывается при помощи следующих терминов:
Очерёдное (последовательное) — листья располагаются по одному (в очередь) на каждый узел.
Супроти́вное — листья располагаются по два на каждом узле и обычно перекрёстно-попарно, то есть каждый последующий узел на стебле развёрнут относительно предыдущего на угол 90°; либо двумя рядами, если не развёрнут, но имеется несколько узлов.
Муто́вчатое — листья располагаются по три и более на каждом особом узле стебля — мутовке. В отличие от супротивных листьев, у мутовчатых каждый последующий завиток может находиться под углом 90° от предыдущего, а может и не находиться, вращаясь на половину угла между листьями в завитке. Следует однако учесть, что супротивные листья могут показаться мутовчатыми на конце стебля.
Розе́точное — листья, расположенные в розетке (пучок листьев, расположенных по кругу из одного общего центра).
Стороны листа
У любого листа в морфологии растений есть две стороны: абаксиальная и адаксиальная.
Абаксиа́льная сторона (от лат. ab — «от» и лат. axis — «ось») — сторона бокового органа побега (листа или спорофилла) растения, обращённая при закладке от конуса нарастания (вершины) побега. Другие названия — спинная сторона, дорзальная сторона.
Противоположная ей сторона называется адаксиа́льной (от лат. ad — «к» и лат. axis — «ось»). Другие названия — брюшная сторона, вентральная сторона.
В подавляющем большинстве случаев абаксиальная сторона — это поверхность листа или спорофилла, обращённая к основанию побега, однако изредка сторона, закладывающаяся абаксиально, разворачивается в процессе развития на 90° или 180° и располагается параллельно продольной оси побега или обращается к его вершине. Это характерно, например, для хвои некоторых видов ели.
Термины «абаксиальный» и «адаксиальный» удобны тем, что позволяют описывать структуры растений, используя само растение как систему отсчёта и не прибегая к двусмысленным обозначениям типа «верхняя» или «нижняя» сторона. Так, для побегов, направленных вертикально вверх, абаксиальная сторона боковых органов будет, как правило, нижней, а адаксиальная — верхней, однако если ориентация побега отклоняется от вертикальной, то термины «верхняя» и «нижняя» сторона могут ввести в заблуждение.
Разделение листовых пластинок
По тому, как листовые пластинки разделены, могут быть описаны две основные формы листьев.
Простой лист состоит из единственной листовой пластинки и одного черешка. Хотя он может состоять из нескольких лопастей, промежутки между этими лопастями не достигают основной жилки листа. Простой лист всегда опадает целиком.
Если выемки по краю простого листа не достигают четверти полуширины листовой пластины, то такой простой лист называется цельным[1].
Сложный лист состоит из нескольких листочков, расположенных на общем черешке (который называется рахис). Листочки, помимо своей листовой пластинки, могут иметь свой черешок (который называется черешочек, или вторичный черешок) и свои прилистники (который называются прилистничками, или вторичными прилистниками). В сложном листе каждая пластинка опадает отдельно. Так как каждый листочек сложного листа можно рассматривать как отдельный лист, при идентификации растения очень важно определить местонахождение черешка. Сложные листья являются характерными для некоторых высших растений, таких как бобовые.
У пальчатых (или лапчатых) листьев все листовые пластинки расходятся по радиусу от окончания корешка подобно пальцам руки. Главный черешок листа отсутствует. Примерами таких листьев может служить конопля (Cannabis) и конский каштан (Aesculus).
У перистых листьев листовые пластинки расположены вдоль основного черешка. В свою очередь, перистые листья могут быть непарноперистыми, с верхушечной листовой пластинкой, например, ясень (Fraxinus); и парноперистыми, без верхушечной пластинки, например, растения из рода Swietenia.
У двуперистых листьев листья разделены дважды: пластинки расположены вдоль вторичных черешков, которые в свою очередь прикреплены к главному черешку; например , альбиция (Albizia).
У трёхлистных листьев имеется только три пластинки; например, клевер (Trifolium), бобовник (Laburnum).
Перстонадрезные листья напоминают перистые, но пластинки у них не полностью разделены; например, некоторые рябины (Sorbus).

Простой лист, Осина (Populus tremula)

Непарноперистый лист, Роза (Rosa)

Пальчатый лист, Конский каштан обыкновенный (Aesculus hippocastanum)

Пальчаторассечённый лист, Морозник чёрный (Helleborus niger)
Характеристики черешков
Черешковые листья имеют черешок — стебелёк, к которому они крепятся. У щитовидных листьев черешок прикреплён внутри от края пластинки. Сидячие и обвивающие листья черешка не имеют. Сидячие листья крепятся прямо к стеблю; у обвивающих листьев листовая пластинка полностью либо частично обволакивает стебель, так что создаётся впечатление, что побег растёт прямо из листа (пример — Клейтония пронзённолистная, Claytonia perfoliata). У некоторых видов акации, например у вида Acacia koa, черешки увеличены и расширены и выполняют функцию листовой пластинки — такие черешки называют филло́дии. На конце филлодия нормальный лист может существовать, а может и нет.
У некоторых растений та часть черешка, которая находится около основания листовой пластинки, утолщена. Это утолщение называется коленце, или геникулум (лат. geniculum). Коленце свойственно, к примеру, для многих растений семейства Ароидные.
Характеристики прилистника
Прили́стник, присутствующий на листьях многих двудольных растений, является придатком на каждой стороне основания черешка и напоминает маленький листик. Прилистники могут опадать по мере роста листа, оставляя после себя рубец; а могут и не опадать, оставаясь вместе с листом (например, так происходит у розовых и бобовых).
Прилистники могут быть:
свободные
сросшиеся — слитые с основанием черешка
раструбовидные — в виде раструба (пример — ревень, Rheum)
опоясывающие основание черешка
межчерешковые, между черешками двух супротивных листьев
межчерешковые, между черешком и противолежащим стеблем
Жилкование
Существует два подкласса жилкования: краевое (основные жилки доходят до концов листьев) и дуговидное (основные жилки проходят почти до концов краёв листа, но поворачивают, не доходя до него).
Типы жилкования:
Сетчатое — локальные жилки расходятся от основных подобно пёрышку и разветвляются на другие маленькие жилки, таким образом создавая сложную систему. Такой тип жилкования типичен для двудольных растений. В свою очередь сетчатое жилкование делится на:
Перисто-нервное жилкование — лист имеет обычно одну основную жилку и множество более мелких, ответвляющихся от основной и идущих параллельно друг к другу. Пример — яблоня (Malus).
Радиальное — лист имеет три основных жилки, исходящих от его основания. Пример — краснокоренник, или цеанотус (Ceanothus).
Пальчатое — несколько основных жилок радиально расходятся недалеко от основания черешка. Пример — клён (Acer).
Параллельное — жилки идут параллельно вдоль всего листа, от его основания до кончика. Типично для однодольных растений, таких как злаки (Poaceae).
Дихотомическое — доминирующие жилки отсутствуют, жилки разделяются на две. Встречается у гинкго (Ginkgo) и некоторых папоротников.
Терминология листа
|
Листья различной формы По часовой стрелке, начиная с правого угла: тройной лопастный, овальный с мелкопильчатым краем, щитовидный с пальчатым жилкованием, заострённый непарноперистый (в центре), перисторассечённый, лопастной, овальный с цельнокрайным краем |

Форма листа
По своей форме лист может быть:
Веерообразный: полукруглый, или в виде веера
Двоякоперистый: каждый листик перистый
Дельтовидный: лист треугольный, крепится к стеблю в основании треугольника
Дланевидный: разделённый на много лопастей
Заострённый: клиновидный с длинной вершиной
Игольчатый: тонкий и острый
Клинообразный: лист треугольный, лист крепится к стеблю на вершине
Копьевидный: острый, с колючками
Ланцетный: лист длинный, широкий посередине
Линейный: лист длинный и очень узкий
Лопастный: с несколькими лопастями
Лопатовидный: лист в виде лопаты
Непарноперистый: перистый лист с верхушечным листиком
Обратноланцетовидный: верхняя часть шире, чем нижняя
Обратносердцевидный: лист в виде сердца, крепится к стеблю на выступающем конце
Обратнояйцевидный: в виде слезы, лист крепится к стеблю на выступающем конце
Овальный: лист овальный, с коротким концом
Овальный: лист овальный, яйцевидный, с заострённым концом в основании
Однолопастный: с одним листиком
Округлый: круглой формы
Пальчатый: лист разделён на пальцевидные лопасти
Парноперистый: перистый лист без верхушечного листика
Перисторассечённый: простой рассечённый лист, у которого сегменты расположены симметрично относительно оси листовой пластины[1]
Перистый: два ряда листиков
Почковидный: лист в форме почки
Рассечённый: листовая пластинка такого листа имеет вырезы, достигающие более двух третей её полуширины; части листовой пластинки рассечённого листа называются сегментами[1]
Ромбовидный: лист в форме ромба
Серповидный: в виде серпа
Сердцевидный: в виде сердца, лист крепится к стеблю в районе ямочки
Стреловидный: лист в виде наконечника стрелы, с расширяющимися лопастями в основании
Триждыперистый: каждый листочек в свою очередь делится на три
Тройчатый: лист разделён на три листочка
Шиловидный: в виде шила
Щитовидный: лист закруглённый, стебель крепится снизу