актиномицетов
колеблется в пределах 0,5—1,5 мк
и
не отличается от диаметра клеток
бактерий, тогда как диаметр нитей грибов
может достигать 50 мк.
В
подавляющем большинстве случаев
мицелий актиномицетов не имеет
перегородок, т.
е. является как бы одной огромной
разветвленной клеткой.
У
высших представителей актиномицетов
(род Actinomyces
по
Н. А. Красильникову) мицелий сохраняется
в течение всей жизни
и
иногда септирован. На плотных
питательных средах такие актиномицеты
образуют более или менее твердые
корковидные или бархатистые колонии
различной величины и окраски, часто
довольно яркой. Часть колонии плотно
врастает в субстрат. На поверхности
имеется воздушный мицелий. Размножение
осуществляется с помощью участков
мицелия и конидий (опор), которые
образуются на концах нитей воздушного
мицелия. Копны нитей со спорами
называются споронос-цавди.
Форма спороносцев (рис. 11) и спор
постоянна для каждого вида и используется
для диагностики актиномицетов.
Споры актиномицетов менее термостойки,
чем бактериальные.

У
менее высокоорганизованных представителей
актиномицетов мицелий относительно
быстро распадается на отдельные
ветвящиеся участки, палочки и даже
кокки, давая очень разнородную
микроскопическую картину. На поверхности
плотных сред такие микроорганизмы
растут в виде довольно плотных колоний,
почти не срастающихся с субстратом.
Воздушный мицелий образуется редко.
Размножение осуществляется фрагментами
мицелия. Конидии (споры) на мицелии не
образуются. В соответствии с определителем
Н. А. Красильникова эта группа объединяется
в род Proactinomyces
(рис.
12). По определителю Берджи часть
представителей этой группы выделена
в род Nocardia.
Наиболее
простыми в морфологическом отношении
являются микобактерии (род Mycobacterium).
Они
имеют рудиментарный мицелий только
на ранних стадиях роста. Обычно в это
время микобактерии имеют вид слегка
разветвленных клеток. Такие разветвленные
клетки очень быстро распадаются на
прямые или слегка искривленные
палочки и кокки (рис. 13). На поверхности
плотных сред микобактерии образуют
мягкие пастообразные или сли-
зистые
колонии разной окраски, не срастающиеся
с субстратом. Конидии и эндоспоры у
микобактерии, по-видимому, не образуются.
По мнению Н. А. Красильникова, многие
микобактерии образуют покоящиеся
клетки — артроспоры. Характерной
особенностью многих микобактерии и
некоторых других представителей актино-
мицетов
является их кислотоустойчивость.
Кислотоустойчивыми называются бактерии,
которые с трудом воспринимают красители.
Клетки таких бактерий окрашиваются
только концентрированными растворами
красителей цри подогревании. Последующее
воздействие кислот не обесцвечивает
клетки микобактерии, и таким образом
они
могут быть дифференцированы от других
бактерий. Кислотоустойчивость клеток
связана с химическим составом их
оболочек. Актиномицеты, как правило,
неподвижны, грамположительны.
Среди
актиномицетов много сапрофитов, но
есть и патогенные виды. Из патогенных
видов наибольшее значение имеют ми-

кобактерии,
вызывающие туберкулез и проказу. Многие
представители
высших актиномицетов имеют огромное
значение
как
продуценты
важнейших антибиотиков — стрептомицина,
хлортетра-циклина
и ряда других.
Актиномицеты
широко распространены в
почвах.
В
воде
они
встречаются
реже.
Это
сравнительно небольшая группа
одноклеточных микроорганизмов,
имеющих спирально извитую форму. Диаметр
клеток спирохет колеблется в пределах
0,25—6 мк,
длина
— от 4 до 500 мк.
Спирохеты
обладают рядом признаков, отличающих
их от других микроорганизмов. Клетки
спирохет не имеют ригидной оболочки,
поэтому они эластичны и_гибки, подобно
клеткам животных. Извитая форма
клеток спирохет поддерживается благодаря
наличию особых фибрилл, которые у
типичных видов состоят из хитиноподобного
вещества — кутина, встречающегося
обычно лишь у животных. Фибриллы могут
быть расположены внутри клетки, но чаще
обнаруживаются вне ее. Фибриллы,
находящиеся
вне клетки, у одних видов образуют так
называемую аксиальную нить, вокруг
которой, как вокруг стержня, спирально
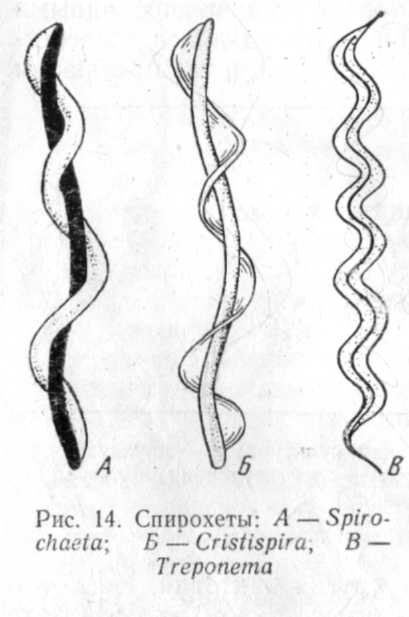
изгибается клетка (рис.4, А,
В),
у
других
— кристу — плоскую килевидную
мембрану
(рис. 4,
Б).
При
разрушении аксиальной нити клетка
спирохеты становится прямой. Напомним,
что в отличие от спирохет, извитая форма
клетки спирилл поддерживается
благодаря наличию ригидной оболочки.
Большинство спирохет подвижны. Они
могут извиваться и сгибаться, а также
перемещаться в жидкой среде путем
вращательных и легких волнообразных
движений. Некоторые сапрофитные формы
могут «ползать» по плотной поверхности,
подобно миксобакте-риям Движение
спирохет связывают с сокращением
фибрилл. Спирохеты, как правило, с
трудом окрашиваются красителями,
которыми хорошо красятся бактериальные
клетки. Их окрашивают методами, которые
применяются для окраски простейших.
Спирохеты размножа-
ются
поперечным изоморфным делением. У
некоторых видов, возможно, существует
цикл размножения, который связан с
образова-нием
небольших
почек или гранул на концах или сбоку
клеток. Иэ
18Спирохеты



каждой
почки или гранулы, по-видимому, может
возникнуть новая клетка спирохеты.
Эндоспоры у спирохет не образуются.
Спирохеты
живут, главным образом, в иловых застойных
водах с низким содержанием кислорода.
Среди спирохет много видов, патогенных
для человека и животных, но есть и
сапрофитные формы. Выращивание спирохет
в чистой культуре налажено недавно,
поэтому физиологические особенности
многих видов этой группы изучены
относительно мало.
В
определителе Берджи спирохеты объединены
в порядок Spirochaetales.
Риккетсии
Риккетсии
являются одноклеточными микроорганизмами,
по некоторым признакам имеющими сходство
как с бактериями, так и с крупными
вирусами. Риккетсии бывают сигаровидной,
палочковидной или овальной формы.
Клетки риккетсии, как и у бактерий,
одеты твердой оболочкой, однако диаметр
клеток составляет всего около 0,1 мк,
а
длина редко превышает 5 мк.
Риккетсии
размножаются, по-видимому, делением,
хотя это точно еще не доказано. Клетки
риккетсии могут быть соединены попарно,
в некоторых случаях они образуют
цепочки, а иногда вытягиваются в довольно
длинные нити. Риккетсии неподвижны, не
образуют эндоспор, грамотрицательны.
Все известные до настоящего времени
представители риккетсии являются
облигатными, обычно внутриклеточными
паразитами и живут в клетках тканей
позвоночных или членистоногих, что
сближает их с вирусами. Ряд риккетсии
вызывает опасные заболевания человека:
сыпной тиф, лихорадку Ку и др. Однако в
отличие от вирусов клетки риккетсии
содержат и ДНК и РНК, а также ряд
ферментов.
Риккетсии
пока удается выращивать только в
культурах тканей или в организме
животных. Их изучением занимаются
медицинские микробиологи.
Сине-зеленые
водоросли
Сине-зеленые
водоросли так же как и бактерии являются
про-кариотными организмами. Однако они
имеют ряд признаков, сближающих их
с растениями.
Среди
сине-зеленых водорослей есть одноклеточные
организмы, имеющие палочковидную
или сферическую форму, размножающиеся
делением и по размерам (род Anacystis)
сходные
с палочковидными бактериями. Иногда
они образуют слизистые скопления.
Большинство сине-зеленых водорослей
— многоклеточные организмы,
представляющие собой длинные, чаще
всего нераз-ветвленные нити. Клетки в
нитях объединены общей наружной стенкой.
Рост нити происходит в результате
одновременного деления клеток,
составляющих нить. Размножение
осуществляется с
19
помощью
гормогоний, отделяющихся с конца нити
или образующихся путем ее распада
на отдельные участки. Некоторым нитчатым
формам (род Anaboena,
Nostoc)
свойственно
образование спор, которые имеют очень
плотную оболочку и способны переживать
неблагоприятные условия. В благоприятных
условиях спора дает новую нить. Многие
сине-зеленые нитчатые водоросли
прикреплены к субстрату. Подвижны
только гормогоний таких водорослей.
Однако среди сине-зеленых водорослей
есть свободноплавающие и даже
подвижные формы. Последние передвигаются
скольжением. Сине-зеленые водоросли
содержат хлорофилл и осуществляют
фотосинтез. Они очень широко распространены
в морских и пресных водах, встречаются
также и в почве. Исследуются сине-зеленые
водоросли главным образом альгологами.
ЭУКАРИОТНЫЕ
МИКРООРГАНИЗМЫ
Грибы
Грибы
— это бесхлорофилльные низшие растения.
Из грибов основным объектом, интересующим
микробиологов, издавна являются
одноклеточные микроскопические грибы
— дрожжи, а также некоторые плесневые
грибы.
Дрожжи.
Дрожжевые
клетки морфологически довольно
разнообразны. Они бывают округлыми,
овальными, удлиненными или
лимоновидными.
Клетки дрожжей значительно крупнее
бактериальных. Диаметр округлых
форм колеблется в среднем от 4 до 12 мк.
Многим
дрожжам свойственно образование более
или менее крупных скоплений различной
формы (рис. 15). Некоторые дрожжи могут
образовывать рудиментарный мицелий.
Оболочка
дрожжевой клетки в отличие от бактериальной
легко различима в обычном световом
микроскопе. Особенно четко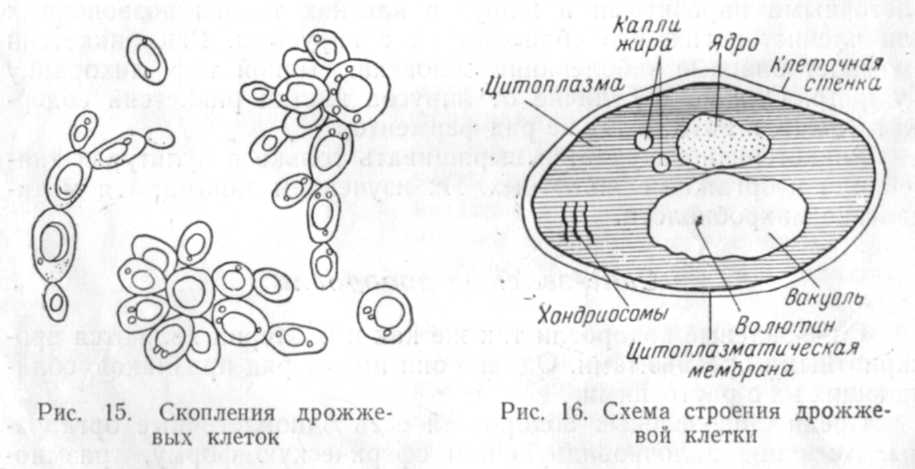





видна
оболочка при исследовании дрожжей с
помощью фазово-контрастного устройства.
В цитоплазме хорошо видны вакуоли. При
специальной окраске в цитоплазме
обнаруживаются дифференцированное
ядро, митохондрии (хондриосомы), гликоген
— запасное вещество типа крахмала,
капельки жира, а в вакуолях — волютин,
основным компонентом которого являются
полифосфаты (рис. 16). Клетки дрожжей
грамположнтельны.
На
плотных питательных средах дрожжи
растут в виде различных выпуклых,
округлых или лопастных, гладких или
складча-
тых,
бесцветных, желтовато-оранжевых или
ярко-розовых, не врастающих в субстрат
колоний мягкой консистенции. Форма и
цвет колоний имеют значение для
диагностики дрожжей.
Размножение
дрожжей осуществляется разными
способами. Большинство дрожжей может
размножаться почкованием (рис. 17, А).
К
почкующимся дрожжам относятся
представители рода Saccharomyces
(сахаромицеты).
Ряд дрожжей размножается делением
(рис. 17, Б).
К
делящимся дрожжам относятся виды рода
Schizosaccharomyces
(шизосахаромицеты).
Кроме того, у многих делящихся и
почкующихся дрожжей наблюдается половой
процесс, связанный со спорообразованием.
Дрожжи, способные к половому процессу,
называются истинными дрожжами и
относятся к классу сумчатых грибов
(аскомицетов). У одних представителей
наблюдается вначале половое слияние
вегетативных клеток, приводящее к
образованию асков (сумок) со спорами
(аскоспора-ми). У других вначале образуются
споры, которые затем копулируют друг
с другом. В каждой дрожжевой клетке или
в аске образуется от двух до восьми,
иногда до 12 спор (рис. 17, В).
Споры
дрожжей имеют толстые оболочки и
достаточно устойчивы к неблагоприятным
воздействиям, хотя и в значительно
меньшей степени, чем эндоспоры
представителей порядка Eubacteriales.
Таким образом, образование спор у
дрожжей — это одновременно и процесс
размножения и формирование устойчивых
форм. Среди дрож-жеподобных организмов
есть аспорогенные, так называемые
ложные дрожжи, которые неспособны
к половому процессу и спорообразованию.
Они относятся к классу несовершенных
грибов.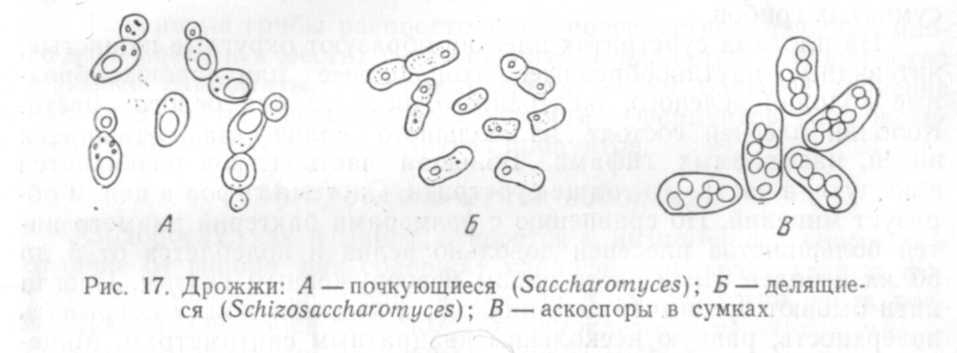




Дрожжи
очень широко распространены в природе.
Они встречаются на винограде, на
других ягодах и фруктах, на овощах, в
молоке, в различных солениях, в воде,
почве, на коже человека и т. д. Многие
дрожжи осуществляют спиртовое брожение
и имеют большое практическое значение.
Они используются для производства
спирта, в хлебопечении, в виноделии,
пивоварении и ряде других отраслей
пищевой промышленности. Среди дрожжей
есть виды, вызывающие заболевания
растений, животных и человека.
Плесневые
грибы.
Под
названием «плесневые грибы» объединяются
некоторые представители фикомицетов,
несовершенных и сумчатых грибов.
На
плотных субстратах плесени образуют
округлые пушистые, нитевидные,
паутинообразные, ватоподобные или
порошкообразные колонии зеленого,
желтоватого, черного или белого цвета.
Колонии плесней состоят из большого
количества ветвящихся нитей, называемых
гифами. Большая часть гифов развивается
в воздухе, а часть — в толще субстрата.
Система гифов в целом образует
мицелий. По сравнению с размерами
бактерий диаметр нитей большинства
плесеней довольно велик и колеблется
от 5 до 50 мк
и
более. Нити часто видны невооруженным
глазом. Иногда нити бывают чрезвычайно
длинными, а мицелий может покрывать
поверхность, равную нескольким квадратным
сантиметрам. Мицелий представителей
фикомицетов не имеет поперечных
перегородок (септ), мицелий других
плесеней септирован. У плесеней
имеется
хорошо дифференцированное ядро, причем
мицелий фикомицетов многоядерный.
Плесени,
как и дрожжи, размножаются бесполым и
половым путем. Оба способа размножения
связаны с образованием спор. Известно
по крайней мере пять способов образования
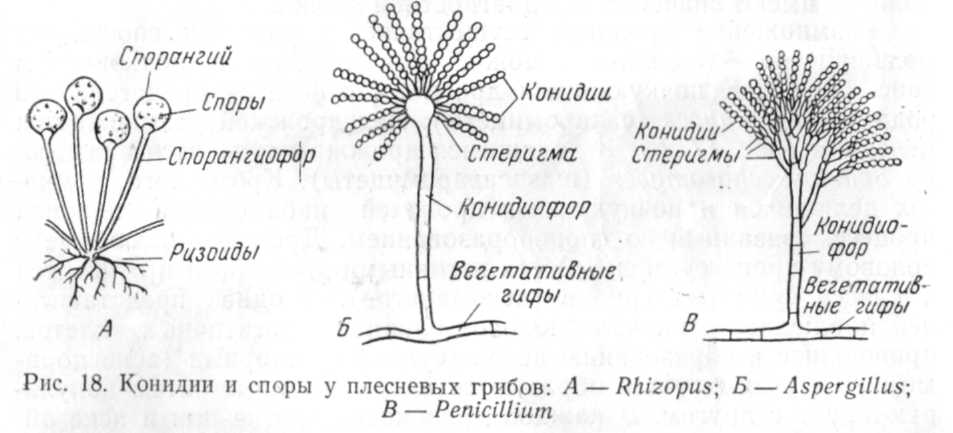
спор бесполым путем. Например, у
фикомицетов споры при бесполом
размножении образуются на свободных
концах плодоносящих гифов — спорангиофорах,
в особых шаровидных тельцах —
спорангиях