

3. Механізм дроблення.
Клітинний цикл соматичних кліток ділиться на чотири стадії (фази). За мітозом (М) відбувається пресинтетичний період (G1), після якого відбувається синтез ДНК (S). Потім наступає премітотична фаза (G2), за якої знову йде мітоз.
Клітинний цикл бластомерів на ранніх стадіях дроблення може бути набагато більш простим - двофазним. Фактори, що регулюють цей двофазний цикл, локалізовані в цитоплазмі. Це ті ж самі фактори, які регулювали розподіли дозрівання при оогенезі, а саме - фактор, що стимулює дозрівання (maturation promoting factor, MPF), цитостатичний фактор (сytostatic factor, CSF) і іони кальцію.
Показано, що в клітинах, які діляться, рівень активності фактор, що стимулює дозрівання, має циклічні зміни. Активність фактору, що стимулює дозрівання, у бластомерах жаби на ранніх стадіях дроблення найвища в М-фазі і не виявляється в S-фазі.
Дія фактору, що стимулює дозрівання, здійснюється шляхом зміни структури ядерної оболонки. Цитостатичний фактор стабілізує фактор дозрівання, затримуючи клітини в стані мітозу, а кальцій інактивує цитостатичний фактор, стимулюючи перехід до S - фази за рахунок інактивації фактору, що стимулює дозрівання. При додаванні цитостатичного фактору припиняються також циклічні скорочення кортикального шару цитоплазми, а наступна ін'єкція іонів кальцію їх стимулює (мал. 3).
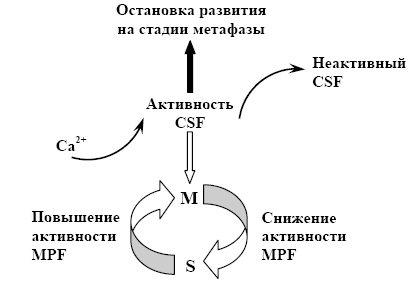
Мал. 3. Модель регуляції клітинного циклу бластомерів (за Гілбертом, 1993)
На ранніх стадіях розвитку цитоплазма визначає швидкість клітинних розподілів і тривалість S- і M-фаз. У період асинхронних розподілів дроблення з'являється фаза G1, подовжується тривалість всіх інших фаз циклу. Починається синтез різних видів РНК на матрицях ДНК, тобто пробуджується транскрипційна активність геному зародка.
Гени, внесені в геном зародка зі сперматозоїдом, проявляють свою дію саме в цей період, у всякому разі, не раніше закінчення періоду синхронного дроблення. Саме в цей час зародок бере свою долю у власні руки і перестає бути генетичною копією матері. Оскільки період асинхронності починається після різного числа розподілів дроблення, то й пробудження транскрипційної активності починається при відповідно різній кількості бластомерів: у ссавців і круглих хробаків практично із самого початку розвитку, в голкошкірих - зі стадії 32 бластомерів, в амфібій - зі стадії бластули.
4. Просторова організація зародка під час дроблення. Борозни дроблення.
Дроблення – суворо координований процес, що знаходиться під генетичним контролем. Видові особливості процесу дроблення визначаються двома основними параметрами:
кількістю та розподілом жовткових білків в цитоплазмі;
присутніми у цитоплазмі факторами, що впливають на орієнтацію мітотичного веретена та часу його утворення.
Якщо жовток у яйці розподілений нерівномірно, то в області, яка містить менше жовтка (анімальна півкуля), клітинні поділи проходять з більшою швидкістю, ніж у протилежній, багатій на жовток області (вегетативна півкуля). Ядро зиготи досить часто буває зміщеним до анімального полюсу.
Закономірності, пов'язані з наявністю та розподілом жовтка в яйцеклітині і напрямком ходу борозен дроблення визначаються 2-мя правилами Гертвіга-Сакса:
- клітинне ядро прагне розташуватися в центрі чистої, вільної від жовтка цитоплазми;
- веретено клітинного розподілу прагне розташуватися по напрямку найбільшого протягу вільної від жовтка цитоплазми.
Найбільш загальною закономірністю голобластичного (див. нижче) дроблення мезо- і оліголецітальних яєць є взаємна перпендикулярність (ортогональність) перших трьох борозен, причому дві перші з них проходять по меридіанах яйця, а третя - по екватору (мал. 4).
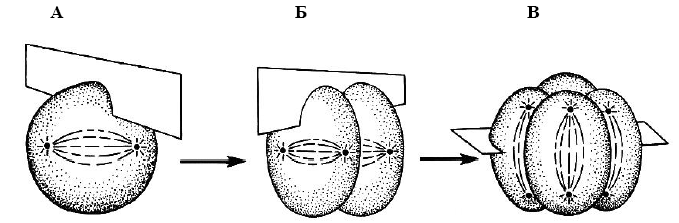
Мал. 4. Хід борозен дроблення (за Гілбертом, 1993):
А – перша меридіональна борозна; Б – друга меридіональна борозна; В – широтна борозна.
Розрізняють борозни дроблення:
- меридіанна (меридіональна) - проходить від анімального полюсу зиготи до вегетативного;
- екваторіальна (її аналогом є широтна борозна, зміщена стосовно екваторіального в напрямку до анімального полюса через перевантаженість жовтком вегетативного полюса) - проходить по екватору зиготи;
- тангенціальна - проходить паралельно поверхні зиготи, у результаті чого утворюється багатошаровий зародок.
Варто відмітити, що швидкість проходження борозен дроблення завжди зворотно пропорційна кількості жовтка в клітині (іноді дане положення називають 3-м правилом дроблення). Дроблення в різних багатоклітинних протікає по-різному. Це залежить від особливостей будови яйцеклітин, насамперед від кількості та розташування в них жовтка, що і лежить в основі класифікації типів дроблення.