

1.3.12. Зв'язки між нервовими клітинами визначають тип поведінки
Поодинокий нейрон людини не дуже відрізняється від поодинокого нейрона черв'яка. Перевага нервової системи людини заснована на величезній кількості клітин, що входять в неї, і. найголовніше, на способі їх з'єднання один з одним, який і визначає можливості нейронів в передачі, комбінуванні і інтерпретації сенсорних сигналів, а також в координації складних послідовностей дій. Щоб зрозуміти клітинні основи еволюції нервової системи, необхідно розглянути механізми, за допомогою яких ембріональні нервові
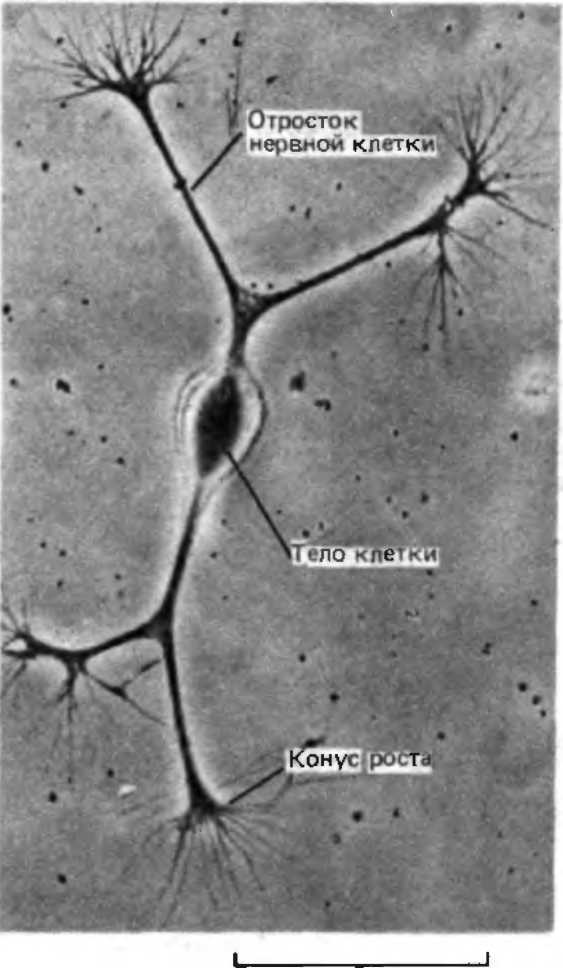
Мал. - Мікрофотографія ізольованої нервової клітини курячого ембріона, поміщеної в культуральну посудину з поживним розчином. У клітини з'являються довгі вирости кожен з яких просувається за допомогою структури, званої конусом зростання.
Клітини придбавають фантастично складні форми і досягають строго впорядкованого способу їх з'єднання (мал. 1-41). Деяке уявлення про дивовижну складність і організованість мереж, які сплітають нервові клітини, що розвиваються, дає зорова система мухи (мал. 1-42). Будова усієї цієї структури задана генетично, і розвиток її відбувається нормально навіть у відсутність світла. При аналізі розвитку нервової системи виявляється, що у усіх видів воно засноване на одних і тих же фундаментальних принципах: однакові механізми використовуються для клітинного руху, міжклітинної адгезії, хімічної сигналізації і так далі. Еволюція складної нервової системи грунтується передусім на еволюції складних контрольних механізмів поєднання і координування цих головних типів клітинної поведінки.
Характер зв'язків між нервовими клітинами визначає поведінка тварини Без жодного навчання і досвіду самець мухи злучається з самицею, павук плете свою павутину, птахи летять на південь. Усі ці дії визначені в ДНК цих видів, яка контролює поведінку окремих клітин при побудові нервової системи у ембріона і роботу нервової системи у дорослого організму.Але не всяка поведінка генетично детермінована, важливе значення має і власний досвід тварини. Позбавлення фоторецепторів, що "дивляться" на одну і ту ж точку, переплетені таким чином, що виявляються приєднаними до одного і тому ж пучка аксонів, що йде в мозок комахи. У кожному оці мухи є присутній більше тисячі таких аксонных пучків, причому кожен з них в ході індивідуального розвитку точно приєднується до правильного набору фоторецепторів. В. Мережа нервових клітин ділянки мозку мухи, одержуючого і оброблювального вхідний сигнал від омматидиев. (А- з люб'язного дозволу Rudi Turner and Anton} Mahowald; У - з N.
Stausfeld, Atlas of an Insect Brain. New York, Springer, 1976.)мозку. Дорослі тварини майже усіх видів - від кишечнополостных до людини - в тій або іншій мірі здатні до навчання. За визначенням навчання - це результат досвіду і, отже, електричній активності нервових клітин, яка повинна призводити до тривалих змін нейронних зв'язків. Висновок Еволюція великих багатоклітинних організмів пов'язана із здатністю эукариотических клітин по-різному экспрессироватъ спадкову інформацію, а також з умінням цих клітин функціонувати спільно. Одним з найбільш ранніх етапів на шляху до многоклеточности була поява епітелію, в якій клітини сполучені в шари, що відділяють внутрішнє середовище організму від зовнішнього оточення. Першими примітивними типами диференційованих клітин Оолжны були бути разом з епітеліальними клітинами нервові клітини м'язові клітини і клітини сполучної тканини. Усі ці типи клітин можна знайти навіть у дуже примітивних сучасних тварин.
Еволюція вищих тварин, використовуючи ту ж основну стратегію розвитку, йшла у напрямі створення усе зростаючого числа спеціалізованих клітинних типів і усе більш витончених методів координації їх активності (мал. 1-43). Дві системи клітин вищих тварин представляють кожна у своєму роді вершину складності багатоклітинної організації. Одна - це імунна система хребетних, клітини якої здатні произвобитъ мільйони різних антитіл. Інша - це нервова система. У нижчих тварин велика частина нейронних зв'язків жорстко генетично детермінована, і програма поведінки еволюціонує лише завдяки мутаціям генетичного матеріалу. У вищих тварин робота і структура нервової системи ставали усе більш схильними до модифікацій (навчанню) завдяки здатності нервових клітин змінювати свої зв'язки у відповідь на викликану зовнішніми стимулами електричну активність.
Клітини використовують чотири основні типи молекул [2]
Певні прості комбінації атомів, такі, як метальные (-СН3), гідроксильні (-ОН), карбоксильні (-СООН) групи і аміногрупи (-№12) - неодноразово повторюються в біологічних молекулах. Кожна така група має певні хімічні і фізичні властивості, які чинять вплив на поведінку будь-яких молекул, що містять такі групи. Загальні відомості про основні типи хімічних груп і їх окремі характерні властивості приведені на схемі 2-2.
Так звані малі органічні молекули клітини є з'єднаннями вуглецю з мол. масою від 100 до 1000, що містять до 30 атомів вуглецю. Молекули такого роду зазвичай знаходяться у вільному стані в цитоплазматичному розчині, утворюючи пул проміжних продуктів, що дають почало великим молекулам, званим макромолекулами. Вони служать також найважливішими проміжними продуктами в хімічних реакціях, що перетворюють витягнуту з їжі енергію в придатну для використання форму (див. нижче).
На долю малих молекул доводиться біля однієї десятої усієї клітинної органічної речовини, причому в клітині є (за наближеними оцінками) присутній близько тисячі різних видів таких молекул. Розщеплюючись, усі біологічні молекули розпадаються до тих простих з'єднань, з яких вони і синтезуються, причому синтез і розпад відбуваються в результаті обмеженої кількості хімічних перетворень, які підкоряються певним правилам. Отже, все наявні в клітині з'єднання .
Лекція 5 Живлення клітин.
Сахара простого типу - моносахариди - є з'єднання із загальною формулою (СН20) п, де п - будь-яке ціле число від трьох до семи. Глюкоза, наприклад, має формулу СлН^Ол (мал. 2-3). Як показано на мал. 2-3. цукри можуть існувати або у формі кільця, або у вигляді відкритого ланцюга. Цукри, що мають структуру відкритого ланцюга, містять гідроксильні групи і, крім того, або альдегідну групу або кетогруппу Альдегідна група і кетогруппа грають особливу роль. По-перше, вони можуть вступити в реакцію з гідроксильною групою тієї ж молекули, що призводить до переходу останньою у форму кільця. Вуглецевий атом початковою альдегідною або кетогруппы можна дізнатися за тією ознакою, що це єдиний в молекулі атом вуглецю, пов'язаний з двома атомами кисню. По-друге, після утворення кільця до цього вуглецевого атома може приєднатися один з атомів вуглецю гідроксильної групи молекули іншого цукру, внаслідок чого утворюється дисахарид (схема 2-3).
Глюкоза служить головним джерелом енергії в багатьох клітинах. В результаті послідовного ряду реакцій окислення (разд. 2.3 2) ця гексоза перетворюється на різні похідні Цукрів з меншою довжиною ланцюга і зрештою розпадається до С02 і Н20. В ході розпаду глюкози вивільняється енергія і генерується відновна здатність, без чого неможливе протікання біосинтетичних реакцій. Енергія, що вивільняється, і генерована відновна сила запасаються у формі двох найважливіших з'єднань - ATP hNADH (см разд. 2.3.1).Прості полісахариди, побудовані із залишків глюкози (у тваринних клітинах це головним чином глікоген, а в рослинних - крохмаль), що повторюються, використовуються для запасання енергії про запас. Проте не можна вважати, що цукри служать виключно для отримання і запасання енергії. Так, з простих полісахаридів складається важливий позаклітинний структурний матеріал (наприклад, целюлоза), а ланцюжки молекул Цукрів, що не повторюються, часто бувають ковалентний пов'язані з білками в глікопротеїнах і з ліпідами в гліколіпідах.2.1.4. Жирні кислоти - компоненти клітинних мембран
У молекулі жирної кислоти, наприклад в пальмітиновій кислоті (мал. 2-5), є дві різні частини: довгий вуглеводневий ланцюг, який має гідрофобний характер (водонерастворима) і хімічно мало активна, і карбоксильна група, що іонізується в розчині, украй гідрофільна (водорозчинна) і така, що легко утворює ефіри і амиды. кислот клітини ковалентний пов'язані з іншими молекулами саме своїми карбоксильними групами. Багато жирних кислот, що виявляються в клітинах, відрізняються один від одного такими хімічними властивостями як довжина вуглеводневого ланцюга, число і положення наявних в них подвійних зв'язків вуглець-вуглець (схема 2-4).
Жирні кислоти є цінним джерелом енергії, оскільки їх розщеплювання супроводжується утворенням такої кількості АТР, яка в два рази перевищує утворення АТР при розщеплюванні такої ж кількості (по масі) глюкози. Жирні кислоти запасаються в цитоплазмі багатьох клітин у вигляді крапельок триацилглицеролов (тригліцеридів). Молекули триацилглицеролов складаються з трьох ланцюгів жирних кислот, кожна з яких приєднана до молекули глицерола (схема 2-4); саме так влаштовані тваринні жири, з якими ми маємо справу в повсякденному житті При необхідності ланцюга жирних кислот можуть вивільнятися з триацилглицеролов і розпадатися до двохвуглецевих одиниць. Такі двохвуглецеві одиниці, що виходять у вигляді ацетогруппы у водорозчинну молекулу, звану ацетил-СоА, піддаються подальшому розщеплюванню в різних экзергонических реакціях, які розглядаються в наступних розділах.
Але найважливіша функція жирних кислот - участь в побудові клітинних мембран. Ці тонкі щільні плівки, якими одягнені усі клітини і вну гриклеточные органели, складаються головним чином з фосфоліпідів - невеликих молекул, схожих з триацилглицеролами наявністю таких компонентів, як жирні кислоти, пов'язані з глицеролом. Проте у фосфоліпідах глицерол частіше пов'язаний не з трьома а лише з двома ланцюгами жирних кислот. Вільне місце, що залишилося, в молекулі глицерола зазвичай займає фосфатна група, яка у свою чергу сполучена з іншими невеликими гідрофільними групами - етаноламіном, холимом або серимом.
У кожної фосфолипидной молекули є гідрофобний хвіст, що складається з ланцюгів двох жирних кислот, і гідрофільна полярна голова, в якій розташовується фосфатна група. Такі фосфолипидные молекули, по суті, є детергенты, про що свідчать їх властивості Невелика кількість фосфоліпідів розподілятиметься по водній поверхні з утворенням фосфолипилного моношару. У такому тонкому шарі дуже тісно зближені хвости обернені у бік повітря, а голови занурені у воду (схема 2-4). Два подібні шари можуть з'єднуватися "хвіст до хвоста", утворюючи в результаті фосфолипидный "сандвіч", або липибный бислой, службовець структурною основою усіх клітинних мембран.
2.1.5. Амінокислоти - субодиниці білківАмінокислоти, що містяться в біологічних тканинах, розрізняються по хімічному складу. Проте усі вони мають ту загальну особливість, що містять карбоксильну групу і аміногрупу, пов'язані з одним і тим же вуглецевим атомом (мал. 2-6). Амінокислоти служать будівельними блоками при синтезі білків - довгих лінійних полімерів амінокислот, сполучених "хвіст до голови" за допомогою пептидного зв'язку між карбоксильною групою однієї амінокислоти і аміногрупою інший (мал. 2-7). У білках зустрічається зазвичай 20 амінокислот з різними бічними ланцюгами, пов'язаними з а-углеродным атомом (схема 2-5). Одні і ті ж 20 амінокислот неодноразово повторюються в усіх білках, у тому числі в білках бактерійного, тваринного і рослинного походження. Можливо, той факт, що саме ці 20 амінокислот було відібрано в ході еволюції, - один з
Мал. 2-8. Заряд бічних ланцюгів амінокислот залежить від значення рН. Карбонові кислоти у водному розчині легко втрачають Н+, утворюючи негативно заряджений іон, що означає за допомогою суфікса "ат", наприклад аспартат або глутамат. Порівнянна ситуація має місце у разі амінів, які, захоплюючи у водному розчині іон Н +, перетворюються на позитивно заряджений іон (він не має спеціальної назви). Ці реакції легко обратимы, і кількість двох форм (зарядженою і незарядженою) залежить від рН розчину. При високих значеннях рН карбонові кислоти мають тенденцію до придбання заряду, а у амінів заряд відсутній. При низьких значеннях рН спостерігається зворотна ситуація: карбонові кислоти не несуть заряду, а аміни заряджені. рН, при якому рівно половина залишків карбонових кислот або амінів заряджена, називають рК амінокислоти. У клітині рН близько до 7 і майже усі карбонові кислоти і аміни знаходяться в зарядженому стані.
Наприклад, у 5 з 20 амінокислот бічні ланцюги можуть нести заряд (мал. 2-8), тоді як бічні ланцюги інших амінокислот электронейтральны, але мають здатність вступати в реакції за допомогою утворення специфічних зв'язків (схема 2-5). Як ми побачимо, особливості бічних ланцюгів усіх амінокислот, що входять до складу цього білку, визначають його властивості і лежать в основі усіх складних і різноманітних функцій білкових молекул.