

Human_Histology
.pdf18 Textbook of Human Histology
Fig. 2.25: Transverse section across a centriole (near its base).
Note nine groups of tubules, each group having three microtubules (Schematic representation)
Fig. 2.26: Comparison of a heterochromatic nucleus (left) and a euchromatic nucleus (right) (Schematic representation)
Fig. 2.27:* ! +/ ! & 3 turns around a complex formed by histones to form a nucleosome.
/ ! 4 ! +/ 3 ! linker-DNA (Schematic representation)
THE NUCLEUS
The nucleus constitute the central, more dense, part of the cell. It is usually rounded or ellipsoid. Occasionally it may be elongated, indented or lobed. It is the largest cell organelle measuring 4–10 μm in diameter. All cells in the body contain nucleus, except mature red blood cells (RBCs) in circulation.
Nuclear Components
Chromatin
Nucleolus
Nuclear membrane
Nucleoplasm.
Chromatin
In usual class-room slides stained with hematoxylin and eosin, the nucleus stains dark purple or blue while the
cytoplasm is usually stained pink. In some cells the nuclei are relatively large and light staining. Such nuclei appear to be made up of a delicate network of fibers—the material making up the fibers of the network is called chromatin (because of its affinity for dyes). At some places the chromatin is seen in the form of irregular dark masses that are called heterochromatin. At other places the network is loose and stains lightly—the chromatin of such areas is referred to as euchromatin. Nuclei which are large and in which relatively large areas of euchromatin can be seen are referred to as open-face nuclei. Nuclei that are made up mainly of heterochromatin are referred to as closedface nuclei (Fig. 2.26).
Nature and Significance of Chromatin
In recent years, there has been a considerable advance in our knowledge of the structure and significance of chromatin. It is made up of a substance called DNA; and of proteins. Most of the proteins in chromatin are histones. Some non-histone proteins are also present.
Each eukaryotic cell contains genetic information encoded in DNA structure. The length of the DNA molecule is 100,000 times longer than nucleus. Therefore, the DNA must be highly folded and tightly packed in the cell nucleus, which is accomplished by the formation of chromatin.
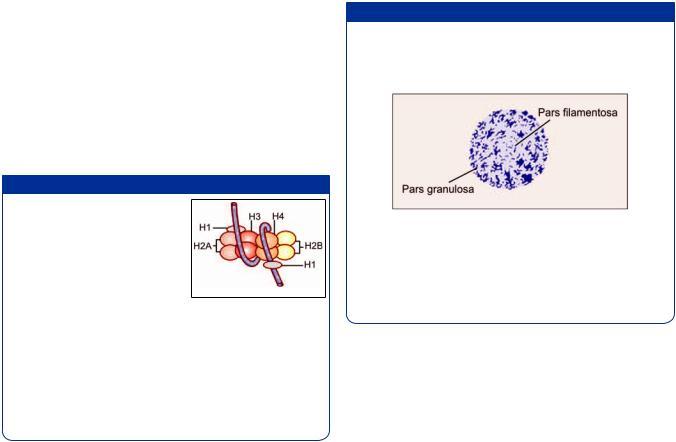
Filaments of DNA form coils around histone complexes. The structure formed by a histone complex and the DNA fiber coiled around it is called a nucleosome. Nucleosomes are attached to one another forming long chains (Fig. 2.27). These chains are coiled on themselves (in a helical manner) to form filaments 30 nm in diameter. These filaments constitute chromatin.
Filaments of chromatin are again coiled on themselves (supercoiling), and this coiling is repeated several times. Each coiling produces a thicker filament. In this way a
filament of DNA that is originally 50 mm long can be reduced to a chromosome only 5 μm in length. (A little calculation will show that this represents a reduction in length of 10,000 times).
Heterochromatin represents areas where chromatin fibers are tightly coiled on themselves forming “solid” masses. In contrast, euchromatin represents areas where coiling is not so marked. During cell division, the entire chromatin within the nucleus becomes very tightly coiled and takes on the appearance of a number of short, thick, rod-like structures called chromosomes. Chromosomes are made up of DNA and proteins. Proteins stabilize the structure of chromosomes.
Added Information
Some details of the formation of a histone complex are shown in Figure 2.28. Five types of histones are recognized. These are H1, H2A, H2B, H3, and H4. Two molecules each of H2A, H2B, H3, and H4jointoformagranularmass,the nucleosome core +/ - ment is wound twice around this core, the whole complex forming a nucleosome. The length of the +/ ! !
contains 146 nucleotide pairs. One nucleosome is connected to the next by a short length of linker DNA. Linker DNA is made up of about 50 nucleotide pairs.
Nucleoli
In addition to the masses of heterochromatin (which are irregular in outline), the nucleus shows one or more rounded, dark staining bodies called nucleoli.
These are spherical and about 1–3 μm in diameter. They stain intensely both with hematoxylin and eosin, the latter giving them a slight reddish tinge. In ordinary preparations, they can be distinguished from heterochromatin by their rounded shape (In contrast masses of heterochromatin are very irregular). Nucleoli are larger and more distinct in cells that are metabolically active.
Using histochemical procedures that distinguish between DNA and RNA, it is seen that the nucleoli have a high RNA content.
Nucleoli are site where ribosomal RNA is synthesized. The templates for this synthesis are located on the related chromosomes. Ribosomal RNA is at first in the form of long fibers that constitute the fibrous zone of nucleoli. It is then broken up into smaller pieces (ribosomal subunits)
Chapter 2 Cell Structure 19
that constitute the granular zone. Finally, this RNA leaves the nucleolus, passes through a nuclear pore, and enters the cytoplasm where it takes part in protein synthesis.
Added Information
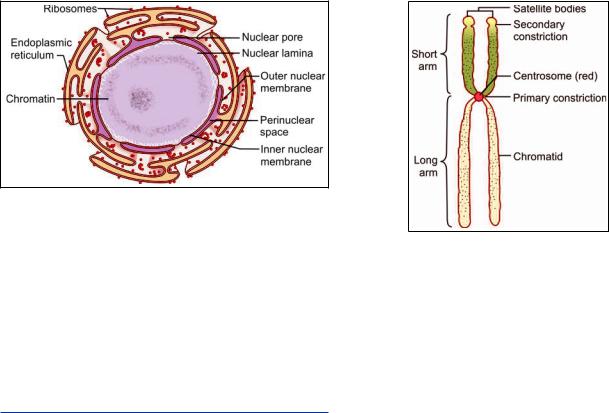
= ># 4 ! zone ( ) and an outer granular zone (pars granulosa) both of which are embedded in an amorphous material (pars amorphosa) (Fig. 2.29).
Fig. 2.29: Electron microscopic structure of a nucleolus
Nucleoli are formed in relationship to the secondary! ! These regions are considered to be nucleolar organizing centers. Parts of the chromosomes located within nucleoli constitute the pars chromosoma of nucleoli.
Nucleoplasm
Besides chromatin and nucleolus, the nucleus also contains various small granules, fibers and vesicles (of obscure function). The spaces between the various constituents of the nucleus are filled by a base called the nucleoplasm.
Nuclear Membrane
With the EM the nucleus is seen to be surrounded by a double-layered nuclear membrane or nuclear envelope. The outer nuclear membrane is continuous with endoplasmic reticulum. The space between the inner and outer membranes is the perinuclear space. This is continuous with the lumen of rough endoplasmic reticulum. The inner layer of the nuclear membrane provides attachment to the ends of chromosomes.
Deep to the inner membrane there is a layer containing proteins and a network of filaments this layer is called the nuclear lamina. Specific proteins present in the inner nuclear membrane give attachment to filamentous proteins of the nuclear lamina. These proteins (called Laminins) form a scaffolding that maintains the spherical shape of the nucleus.
At several points the inner and outer layers of the nuclear membrane fuse leaving gaps called nuclear pores. Each pore is surrounded by dense protein arranged in the form of eight complexes. These proteins and the pore together form the pore complex.
20 Textbook of Human Histology
Fig. 2.30: Nuclear envelope (Schematic representation)
Nuclear pores represent sites at which substances can pass from the nucleus to the cytoplasm and vice versa (Fig. 2.30). The nuclear pore is about 80 nm across. It is partly covered by a diaphragm that allows passage only to particles less than 9 nm in diameter. A typical nucleus has 3,000–4,000 pores.
It is believed that pore complexes actively transport some proteins into the nucleus, and ribosomes out of the nucleus.
CHROMOSOMES
Haploid and Diploid Chromosomes
During cell division, the chromatin network in the nucleus becomes condensed into a number of thread-like or rod-like structures called chromosomes. The number of chromosomes in each cell is fixed for a given species, and in man it is 46. This is referred to as the diploid number (diploid = double). However, in spermatozoa and in ova the number is only half the diploid number, i.e. 23—this is called the haploid number (haploid = half).
Autosomes and Sex Chromosomes
The 46 chromosomes in each cell can again be divided into 44 autosomes and two sex chromosomes. The sex chromosomes may be of two kinds, X or Y. In a man, there are 44 autosomes, one X chromosome and one Y chromosome; while in a woman, there are 44 autosomes and two X chromosomes in each cell. When we study the 44 autosomes we find that they really consist of 22 pairs, the two chromosomes forming a pair being exactly alike (homologous chromosomes). In a woman, the two X chromosomes form another such pair; but in a man this pair is represented by one X and one Y chromosome. One chromosome of each pair is obtained (by each individual) from the mother, and one from the father.
Fig. 2.31: A typical chromosome. Note that this chromosome is submetacentric (Schematic representation)
As the two sex chromosomes of a female are similar the female sex is described as homogametic; in contrast the male sex is heterogametic.
Structure of Fully Formed Chromosomes
Each chromosome consists of two parallel rod-like elements that are called chromatids (Fig. 2.31). The two chromatids are joined to each other at a narrow area that is light staining and is called the centromere (or kinetochore). In this region the chromatin of each chromatid is most highly coiled and, therefore, appears to be thinnest. The chromatids appear to be “constricted” here and this region is called the primary constriction.
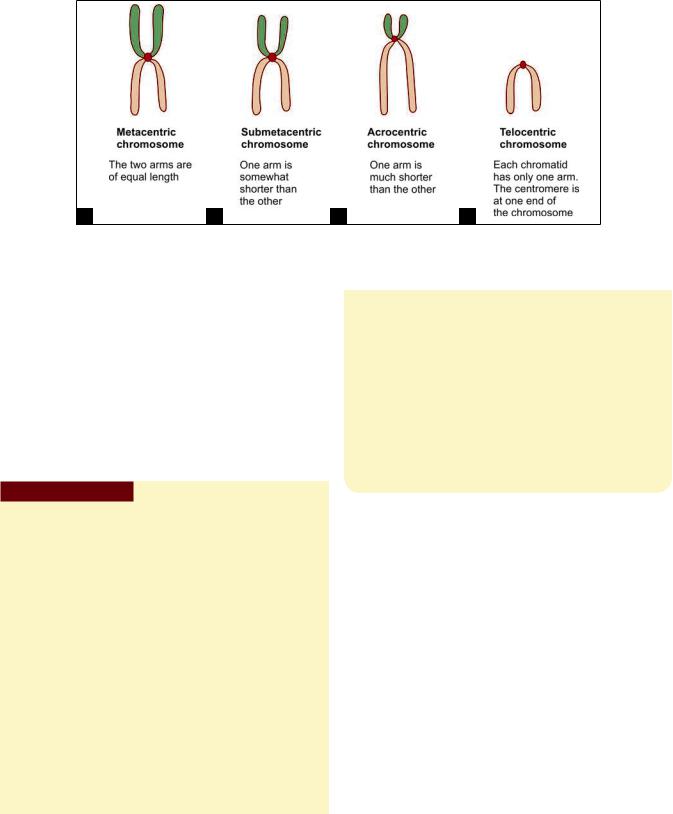
Typically the centromere is not midway between the two ends of the chromatids, but somewhat towards one end. As a result each chromatid can be said to have a long arm and a short arm. Such chromosomes are described as being submetacentric (when the two arms are only slightly different in length); or as acrocentric (when the difference is marked) (Figs. 2.32A to D). In some chromosomes, the two arms are of equal length such chromosomes are described as metacentric. Finally, in some chromosomes the centromere may lie at one end, such a chromosome is described as telocentric.
Differences in the total length of chromosomes, and in the position of the centromere are important factors in distinguishing individual chromosomes from each other. Additional help in identification is obtained by the presence in some chromosomes of secondary constrictions. Such constrictions lie near one end of the chromatid. The part of the chromatid “distal” to the constriction may appear to be a rounded body almost separate from the rest of the chromatid such regions are called satellite
Chapter 2 Cell Structure 21
A B C D
Figs. 2.32A to D: Nomenclature used for different types of chromosomes, based on differences in lengths of the two arms of each chromatid (Schematic representation)
bodies (Secondary constrictions are concerned with the formation of nucleoli and are, therefore, called nucleolar organizing centers). Considerable help in identification of individual chromosomes is also obtained by the use of special staining procedures by which each chromatid can be seen to consist of a number of dark and light staining transverse bands.
Chromosomes are distinguishable only during mitosis. In the interphase (between successive mitoses) the chromosomes elongate and assume the form of long threads. These threads are called chromonemata (singular = chromonema).
Pathological Correlation
Karyotyping
Using the criteria described above, it is now possible to identify each chromosome individually and to map out the chromosomes of an individual. This procedure is called karyotyping. For this purpose a sample of blood from the individual is put into a suitable medium in which lymphocytes can multiply. After a few hours a drug (colchicine, colcemid) that arrests cell division at a stage when chromosomes are most distinct, is added to the medium. The dividing cells are then treated with hypotonic saline so that they swell up. This facilitates the proper spreading out of chromosomes. A suspension containing the dividing cells is spread out on a slide and suitably stained. Cells in which the chromosomes are well spread out (without overlap) are photographed. The photographs are cut-up and the chromosomes arranged in proper sequence. In this way a map of chromosomes is obtained, and abnormalities in their number or form can be!
(For details of chromosomal abnormalities see the author’s HUMAN EMBRYOLOGY).
Contd...
Contd...
In recent years greater accuracy in karyotyping has been achieved by use of several different banding techniques, and by use of computerized analysis.
It has been estimated that the total DNA content of a cell (in all chromosomes put together) is represented by about 6 × 109 nucleotide pairs. Of these 2.5 × 108 are present in chromosome 1 (which is the largest chromosome). The Y-chromosome (which is the smallest chromosome) contains 5 × 107 nucleotide pairs.
In the region of the centromere, the DNA molecule is specialized for attachment to the spindle. This region is surrounded by proteins that form a mass. This mass is the kinetochore. The ends of each DNA molecule are also specialized. They are called telomeres.
Significance of Chromosomes
Each cell of the body contains within itself a store of information that has been inherited from precursor cells. This information (which is necessary for the proper functioning of the cell) is stored in chromatin. Each chromosome bears on itself a very large number of functional segments that are called genes. Genes represent “units” of stored information which guide the performance of particular cellular functions, which may in turn lead to the development of particular features of an individual or of a species. Recent researches have told us a great deal about the way in which chromosomes, and genes store and use information.
The nature and functions of a cell depend on the proteins synthesized by it. Proteins are the most important constituents of our body. They make up the greater part of each cell and of intercellular substances. Enzymes, hormones, and antibodies are also proteins.
It is, therefore, not surprising that one cell differs from another because of the differences in the proteins
22 Textbook of Human Histology
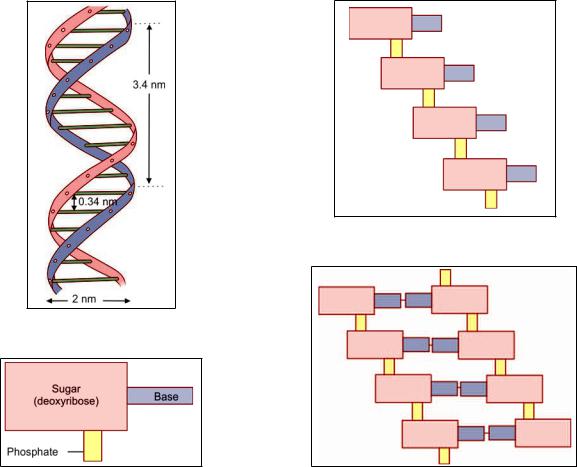
Fig. 2.33: Part of a DNA molecule arranged in the form of a double helix (Schematic representation)
Fig. 2.34: Composition of a nucleotide. The base may be adenine, cytosine, guanine or thymine (Schematic representation)
Fig. 2.35: Linkage of nucleotides to form one strand of a DNA molecule (Schematic representation)
Fig. 2.36: Linkage of two chains of nucleotides to form part of a DNA molecule (Schematic representation)
that constitute it. Individuals and species also owe their distinctive characters to their proteins. We now know that chromosomes control the development and functioning of cells by determining what type of proteins will be synthesized within them.
Chromosomes are made up predominantly of a nucleic acid called DNA, and all information is stored in molecules of this substance. When the need arises this information is used to direct the activities of the cell by synthesizing appropriate proteins. To understand how this becomes possible we must consider the structure of DNA in some detail.
Basic Structure of DNA
DNA in a chromosome is in the form of very fine fibers. If we look at one such fiber, it has the appearance as shown in Figure 2.33. It is seen that each fiber consists of two strands that are twisted spirally to form what is called a double helix. The two strands are linked to each other at regular intervals (note the dimensions shown in Fig. 2.33).
Each strand of the DNA fiber consists of a chain of nucleotides. Each nucleotide consists of a sugar deoxyribose, a molecule of phosphate and a base (Fig. 2.34). The phosphate of one nucleotide is linked to the sugar of the next nucleotide (Fig. 2.35). The base that is attached to the sugar molecule may be adenine, guanine, cytosine, or thymine. The two strands of a DNA fiber are joined together by the linkage of a base on one strand with a base on the opposite strand (Fig. 2.36).
This linkage is peculiar in that adenine on one strand is always linked to thymine on the other strand, while cytosine is always linked to guanine. Thus the two strands are complementary and the arrangement of bases on one strand can be predicted from the other.
The order in which these four bases are arranged along the length of a strand of DNA determines the nature of the protein that can be synthesized under its influence. Every protein is made up of a series of amino acids; the nature of the protein depending upon the amino acids present, and the sequence in which they are arranged. Amino acids
Chapter 2 Cell Structure 23
may be obtained from food or may be synthesized within the cell. Under the influence of DNA these amino acids are linked together in a particular sequence to form proteins.
Added Information
In the preceding paragraphs the structure of DNA has been described in the simplest possible terms. We will now consider some details:
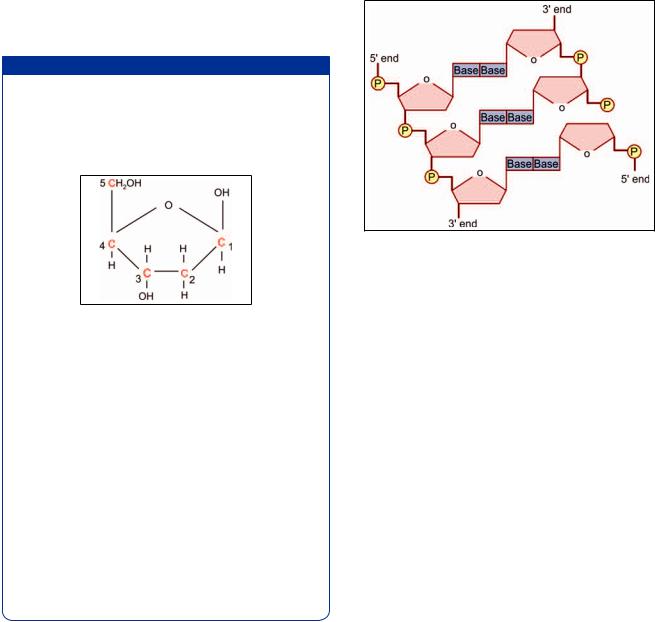
The structure of the sugar deoxyribose is shown in? / 4 ! @ also note how they are numbered.
Fig. 2.37: Structure of deoxyribose. Note the numbering of carbon atoms (Schematic representation)
Next observe, in Figure 2.38, that C-3 of one sugar molecule is linked to C-5 of the next molecule through a phosphate linkage (P). It follows that each strand of DNA has a 5’ end and a 3’ end.
Next observe that although the two chains forming DNA are similar they are arranged in opposite directions. In Figure 2.38, the 5’ end of the left chain, and the 3’ The two chains of nucleotides are, therefore, said to be antiparallel.
The C-1 carbon of deoxyribose give attachment to a base. This base is attached to a base of the opposite chain as already described.
The reason why adenine on one strand is always linked to thymine on the other strand is that the structure of these two molecules is complementary and hydrogen bonds are easily formed between them. The same is true for cytosine and guanine.
Ribonucleic Acid
In addition to DNA, cells contain another important nucleic acid called ribonucleic acid or RNA. The structure of a molecule of RNA corresponds fairly closely to that of one strand of a DNA molecule, with the following important differences:
RNA contains the sugar ribose instead of deoxyribose.
Instead of the base thymine it contains uracil.
RNA is present both in the nucleus and in the cytoplasm
of a cell. It is present in three main forms namely messenger RNA (mRNA), transfer RNA (tRNA), and ribosomal RNA (rRNA). Messenger RNA acts as an intermediary between
Fig. 2.38: Nucleotides are linked to form a chain of DNA. The asymmetric placing of bonds gives a helical shape to the chain (Schematic representation)
the DNA of the chromosome and the amino acids present in the cytoplasm and plays a vital role in the synthesis of proteins from amino acids.
Some forms of RNA are confined to nuclei. The small nuclear RNAs (SnRNA) are concerned with RNA splicing.
Synthesis of Protein
A protein is made up of amino acids that are linked together in a definite sequence. This sequence is determined by the order in which the bases are arranged in a strand of DNA. Each amino acid is represented in the DNA molecule by a sequence of three bases (triplet code). It has been mentioned earlier that there are four bases in all in DNA, namely adenine, cytosine, thymine, and guanine. These are like letters in a word. They can be arranged in various combinations so that as many as sixtyfour “code words” can be formed from these four bases. There are only about twenty amino acids that have to be coded for so that each amino acid has more than one code. The code for a complete polypeptide chain is formed when the codes for its constituent amino acids are arranged in proper sequence. That part of the DNA molecule that bears the code for a complete polypeptide chain constitutes a structural gene or cistron.
At this stage it must be emphasized that a chromosome is very long and thread-like. Only short lengths of the fiber are involved in protein synthesis at a particular time.
The main steps in the synthesis of a protein may now be summarized as follows (Fig. 2.39):
The two strands of a DNA fiber separate from each other (over the area bearing a particular cistron) so that the ends of the bases that were linked to the opposite strand are now free.
24 Textbook of Human Histology
Fig. 2.39: J K +/ The process is actually more complex as explained in the text
(Schematic representation)
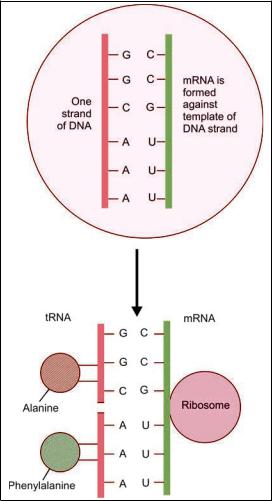
A molecule of mRNA is synthesized using one DNA strand as a guide (or template), in such a way that
one guanine base is formed opposite each cytosine base of the DNA strand, cytosine is formed opposite guanine, adenine is formed opposite thymine, and uracil is formed opposite adenine. In this way the code for the sequence in which amino acids are to be linked is passed on from DNA of the chromosome to mRNA. This process is called transcription. (Transcription takes place under the influence of the enzyme RNA polymerase).
That part of the mRNA strand that bears the code for one amino acid is called a codon.
This molecule of mRNA now separates from the DNA strand and moves from the nucleus to the cytoplasm (passing through a nuclear pore).
In the cytoplasm, the mRNA becomes attached to a ribosome.
As mentioned earlier, the cytoplasm also contains another form of RNA called tRNA. In fact there are about twenty different types of tRNA each corresponding to one amino acid. On one side tRNA becomes attached to an amino acid. On the other side it bears a code of three bases (anticodon) that are complementary to the bases coding for its amino acid on mRNA. Under the influence of the ribosome several units of tRNA, along with their amino acids, become arranged along side the strand of mRNA in the sequence determined by the code on mRNA. This process is called translation.
The amino acids now become linked to each other to form a polypeptide chain. The amino acids are linked up exactly in the order in which their codes are arranged on mRNA, which in turn is based on the code on the DNA molecule. Chains of amino acids formed in this way constitute polypeptide chains. Proteins are formed by union of polypeptide chains.
The flow of information from DNA to RNA and finally
to protein has been described as the “central dogma of molecular biology”.
Epithelia
One or more layers of cells that cover the outer surface (of the body) or line the luminal surface of tubular structures and cavities of the body are called epithelia (singular = epithelium).
CHARACTERISTIC FEATURES OF
EPITHELIAL TISSUE
Very cellular with little intercellular space (20 nm)
Usually avascular
Cells rest on a basement membrane
Cells show polarity
Cells may display surface modifications
FUNCTIONS
Protection
Absorption
Secretion
Exchange
CLASSIFICATION OF EPITHELIA (FLOWCHART 3.1)
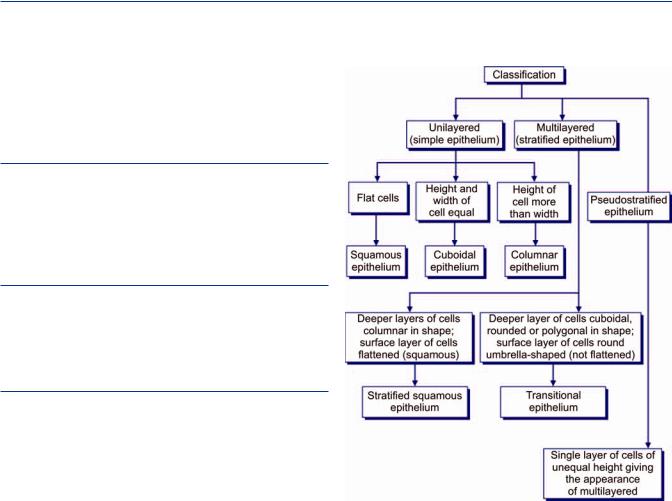
An epithelium may consist of only one layer of cells when it is called a unilayered or simple epithelium. Alternatively, it may be multilayered (stratified) or it can be pseudostratified.
Unilayered (simple) epithelia: Single layer of cells resting on a basement membrane. It may be further classified according to the shape of the cells constituting them.
When the cells are flattened, their height being very little as compared to their width. Such an epithelium is called as a squamous epithelium.
When the height and width of the cells of the epithelium are more or less equal (i.e. they look like squares in section) it is described as a cuboidal epithelium.
When the height of the cells of the epithelium is distinctly greater than their width, it is described as a columnar epithelium.
Flowchart 3.1:
Pseudostratifiedcolumnarepithelia: In true sense this is a simple epithelium as each cell rests on the basement membrane. This epithelium gives an appearance of a multilayered epithelium due to unequal height and shape of cells.
Multilayered (stratified) epithelia: Epithelia which consist of multiple layers with the basal layer resting on the basement membrane. The epithelium is named according to the shape of cells of the most superficial layer.
26 Textbook of Human Histology
PLATE 3.1: Squamous Epithelium
A
B C D
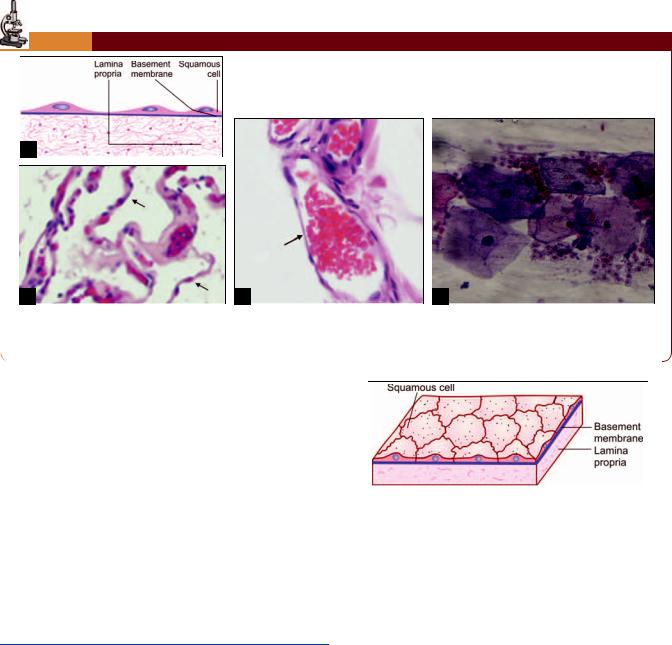
Simple squamous epithelium. (A) As seen in drawing; (B) An alveolus of the lung showing a lining of simple squamous epithelium (photomicrograph) (see arrows); (C) A capillary lined by endothelium (photomicrograph) (see arrow);
(D) Surface view as seen in buccal smear (photomicrograph)
Stratified squamous: The deeper layers are columnar, but in proceeding toward the surface of the epithelium the cells become increasingly flattened (or squamous). It may be noted that all cells in this kind of epithelium are not squamous.
Stratified cuboidal: The surface cells are cuboidal in shape.
Stratified columnar: The surface cells are columnar in shape.
Transitional epithelium: In this type of multilayered epithelium all layers are made up of cuboidal, polygonal, or round cells. The cells toward the surface of the epithelium are round. As transitional epithelium is confinedtotheurinarytract,itisalsocalledurothelium.
SIMPLE EPITHELIUM
Squamous Epithelium
Description
In surface view, the cells have polygonal outlines that interlock with those of adjoining cells (Plate 3.1D ).
In a section, the cells appear flattened their height being much less as compared to their width (Fig. 3.1).
The cytoplasm of cells forms only a thin layer. The nuclei produce bulgings of the cell surface (Plate 3.1).
With the electron microscope (EM) the junctions between cells are marked by occluding junctions. The
Fig. 3.1:
junctions are thus tightly sealed and any substance passing through the epithelium has to pass through the cells, and not between them.
Location
Squamous epithelium lines the alveoli of the lungs.
It lines the free surface of the serous pericardium, the pleura, and the peritoneum; here it is called mesothelium.
It lines the inside of the heart, where it is called endocardium; and of blood vessels and lymphatics, where it is called endothelium.
Squamous epithelium is also found lining some parts of the renal tubules, and in some parts of the internal ear.
Function
It helps in rapid transport of substances, diffusion of gases and filtration of fluids.
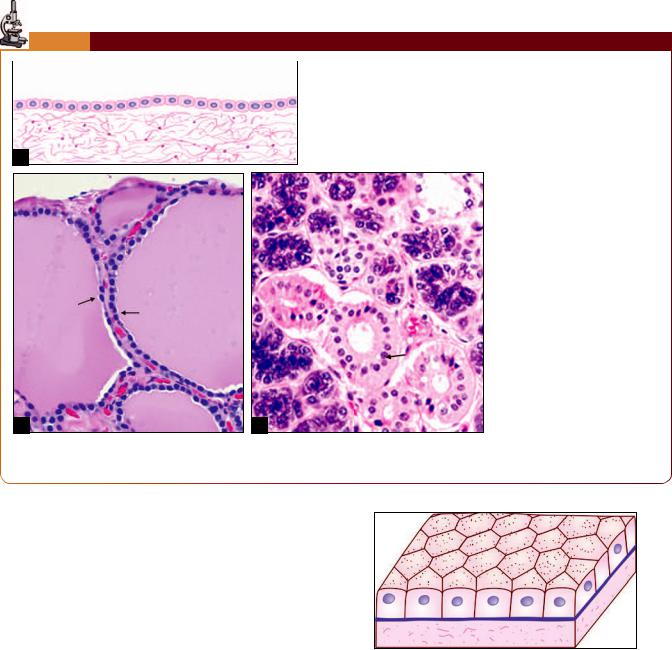
Chapter 3 Epithelia 27
PLATE 3.2: Cuboidal Epithelium
! "
! #$ |
|
% |
& |
|
|
A
B C
Simple cuboidal epithelium. (A) As seen in a section (drawing);
(B)Thyroid follicle lined by simple cuboidal epithelium (photomicrograph) (see arrows);
(C)Duct of salivary gland (photomicrograph) (see arrow)
Cuboidal Epithelium
Description
In cuboidal epithelium, the height of the cells is about the same as their width. The nuclei are usually rounded (Plate 3.2).
In sectional view cells appear cuboidal in shape. When viewed from surface, cells are hexagonal in shape (Fig. 3.2).
Location
A typical cuboidal epithelium may be seen in the follicles of the thyroid gland, in the ducts of many glands, and on the surface of the ovary.
Other sites are the choroid plexuses, the inner surface of the lens, and the pigment cell layer of the retina.
A cuboidal epithelium with a prominent brush border is seen in the proximal convoluted tubules of the kidneys.
Function
It is mainly concerned with secretory and absorptive functions.
Fig. 3.2:
Columnar Epithelium
Description
Cells of the epithelium are much taller compared to their width. Nuclei are elongated and located in the lower half of the cells. All nuclei are placed at the same level in neighboring cells (Fig. 3.3 and Plate 3.3).