

Глава 5 95

Рис. 5.7. Положение спирометра после глубокого выдоха.
1 — внутренний цилиндр; 2 — внешний цилиндр
После первого вдоха ребенка, когда легкие наполняются воздухом, и до конца жизни при любых условиях легкие полностью не освобождаются от воздуха. Даже после максимального выдоха в легких остается около 1000—1500мл воздуха. Этот объем воздуха получил название остаточного воздуха. Остаточный воздух сохраняется даже в легких трупа. Это видно из того, что если бросить в воду кусок легкого мертворожденного ребенка, то он потонет, а если бросить кусок легкого трупа взрослого человека или ребенка, дышавшего хотя бы короткое время, то он не потонет, а будет плавать на поверхности воды. Как известно, газообмен происходит только в альвеолах; воздух же, находящийся в воздухоносных путях, в газообмене участия не принимает. Таким образом, не весь вдыхаемый воздух участвует в газообмене, а только та часть, которая попадает в альвеолы. При обычном дыхании мы вдыхаем 500 мл, из которых около 140 мл остаются в воздухоносных путях, а около 360 мл поступают в альвеолы. Эти 140 мл задерживаются в гортани, трахее, бронхах и бронхиолах и во время дыхания изменениям не подвергаются. Пространство, заполненное воздухом, не участвующим в газообмене, называется вредным пространством.
Параметры легочной вентиляции должны соответствовать потребности организма в кислороде в каждый момент времени. Параметры дыхания изменяются под влиянием многих факторов. Например, при высокой физической нагрузке происходит повышенное потребление кислорода и требуется, соответственно, интенсификация легочной вентиляции, которая достигается за счет изменения глубины и частоты дыхания. Для специалиста-полиграфолога важно знать, что определенные эмоции также нередко вызывают кратковременные изменения параметров дыхательного процесса. Регуляция снабжения организма кислородом и удаление из организма углекислого газа осуществляются скоординированной работой нервной, дыхательной, сердечно-сосудистой систем и крови. Общая схема регуляции процесса дыхания приведена на рис. 5.8.
В регуляции дыхания принимают участие как рефлекторные, так и гуморальные механизмы. Ритм дыхания обусловлен тем, что к дыхательной мускулатуре (межреберные мышцы, диафрагма и др.) из центральной нервной системы беспрерывно поступают импульсы, вызывающие ее сокращение. Еще в прошлом веке экспериментально было доказано, что возбуждение, вызывающее сокращение дыхательных мышц, возникает в продолговатом мозге. Эти опыты заключались в том, что производилась перерезка продолговатого мозга на разных уровнях. Оказалось, что перерезка на уровне верхней границы продолговатого мозга не вызывает значительных нарушений дыхания, а перерезка между продолговатым и спинным мозгом приводит к полной остановке дыхания. Аналогичные результаты были получены при охлаждении этих же участков центральной нервной системы. Охлаждение верхней границы продолговатого мозга не вызывает прекращения дыхания, между тем как охлаждение участка на границе продолговатого и спинного мозга
Барорецепторы
и хеморецепторы
в дуге аорты и каротидном
синусе
Пневмотоксические
центры в
отделах мозга
Дыхательные
центры продолговатого мозга
Межреберные
/мышцы
Ребра
Кожа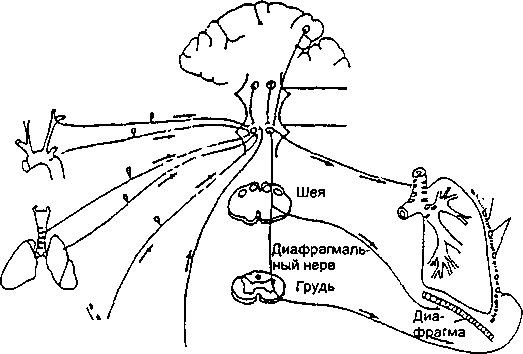
![]() Межреберные. Л—^Ребра
Межреберные. Л—^Ребра
мышцы к/
t rw- \
, i-natijt *Яа»Л
Диафрагма АФФЕРЕНТНЫЕ ПУТИ
ЭФФЕРЕНТНЫЕ ПУТИРис. 5.8. Схема регуляции процесса дыхания
влечет за собой полное прекращение дыхания. Группа нейронов, деятельность которых обусловливает сокращение дыхательной мускулатуры, получила название дыхательного центра. Дыхательный центр является двусторонним, и при нарушении какой-либо его половины прекращается сокращение дыхательной мускулатуры этой же стороны. Разрушение обеих половин дыхательного центра вызывает полную остановку дыхания. Путем электрического раздражения отдельных точек продолговатого мозга удалось доказать существование двух групп нейронов (рис. 5.9), а именно нейронов вдоха (инспираторных) и нейронов выдоха (экспираторных).
Корешки IX, Хч.м.н.
Корешек С1
Рис. 5.9. Расположение инспираторных (И) и экспираторных (Э) нейронов в области продолговатого мозга
Э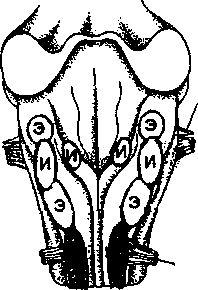 ти
нейроны обеспечивают ритмичность вдоха
и выдоха. Дыхательный
центр в продолговатом мозгу обладает
свойством автоматии. Это означает, что
его нейроны продолжают ритмично
генерировать импульсы даже в отсутствие
нервных импульсов, поступающих по
центростремительным путям. В дальнейшем
стало очевидно, что дыхание хотя и
возможно только при целости дыхательного
центра, находящегося в продолговатом
мозгу,
но если отделить продолговатый мозг, а
следовательно, и дыхательный
центр от высших отделов головного мозга,
то регуляция
дыхания нарушается. Исследования
позволили обнаружить
в стволовых структурах целых шесть
типов (рис. 5.10) нейронов,
возбуждение которых связано с различными
фазами дыхания.
В респираторном цикле помимо фазы
инспирации была также отдельно выделена
фаза постинспирации.
Постинспирация
связана с тем, что на максимуме вдоха
сила растяжения легкого
является очень высокой, а потому выдох
сначала идет медленно, а инспираторная
мускулатура остается в это время в
сокращенном состоянии. Эта промежуточная
фаза дыхательного цикла и получила
название постинспирации.
ти
нейроны обеспечивают ритмичность вдоха
и выдоха. Дыхательный
центр в продолговатом мозгу обладает
свойством автоматии. Это означает, что
его нейроны продолжают ритмично
генерировать импульсы даже в отсутствие
нервных импульсов, поступающих по
центростремительным путям. В дальнейшем
стало очевидно, что дыхание хотя и
возможно только при целости дыхательного
центра, находящегося в продолговатом
мозгу,
но если отделить продолговатый мозг, а
следовательно, и дыхательный
центр от высших отделов головного мозга,
то регуляция
дыхания нарушается. Исследования
позволили обнаружить
в стволовых структурах целых шесть
типов (рис. 5.10) нейронов,
возбуждение которых связано с различными
фазами дыхания.
В респираторном цикле помимо фазы
инспирации была также отдельно выделена
фаза постинспирации.
Постинспирация
связана с тем, что на максимуме вдоха
сила растяжения легкого
является очень высокой, а потому выдох
сначала идет медленно, а инспираторная
мускулатура остается в это время в
сокращенном состоянии. Эта промежуточная
фаза дыхательного цикла и получила
название постинспирации.
Возбуждение нейронов отображено более светлым оттенком серого цвета; торможение нейронов изображено темным оттенком серого цвета.
Последующим изучением было доказано непосредственное участие коры головного мозга в регуляции дыхания. Участие коры больших полушарий в регуляции дыхания определяется и тем, что в течение жизни образуется множество условных рефлексов на дыхание. Таким образом, «гра-
Рис. 5.10. Активность
различных типов дыхательных
мотонейронов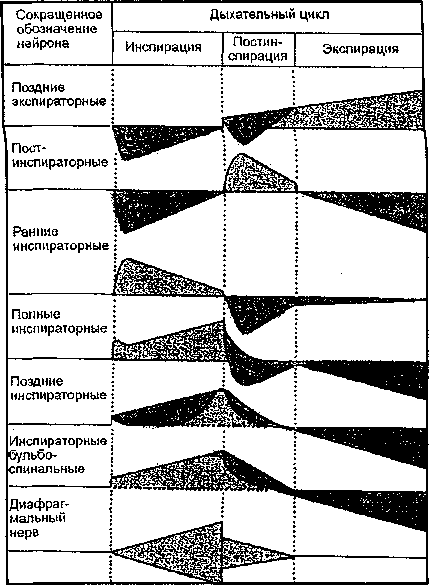
Рис.
5.11. Место разветвления общей сонной
артерии на внешнюю и внутреннюю
сонные артерии:
1
— общая
сонная артерия; 2
—
каротидный
синус;
3
—
внутренняя
сонная артерия; 4
—
внешняя
сонная артерия; 5
— синокаротид-
ный
нерв; 6
— языкоглоточный
нерв
Рефлекторное изменение дыхания можно наблюдать при раздражении центростремительных нервов других участков нашего тела. Например, общеизвестно, что во время купания при опускании ног в холодную воду происходит кратковременная остановка дыхания в состоянии вдоха. Такая остановка является результатом рефлекторного влияния на дыхательный центр.
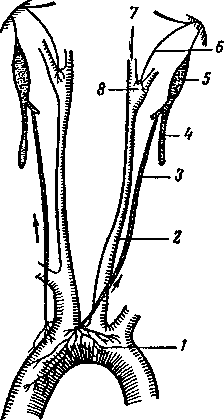
Большое значение имеют рефлексы с сосудистых рефлексогенных зон, например с хеморецепторов каротидного синуса2, который находится у места разветвления общей сонной артерии на внешнюю и внутреннюю сонные артерии (рис. 5.11). Хеморецепторы, ипульсация с которых поступает в дыхательный центр, расположены также в области дуги аорты (рис. 5.12).
Повышение кровяного давления раздражает окончания каротидного и депрессорного нервов. Раздражение этих окончаний вызывает падение кровяного давления. Одновременно отсюда же возникает рефлекторное влияние и на дыхательный центр. Повышение давления крови вызывает задержку дыхания, а понижение давления — усиление дыхания. Как мы уже сказали выше, рецепторы каротидного синуса чувствительны именно к действию гипоксического фактора (недостатка кислорода в крови).
Рис. 5.12. Схема
депрессорного нерва
аорты и сонной
артерии:
1 — дуга аорты; 2 — общая сонная артерия; 3 — депрес-сорный нерв; 4 — блуждающий нерв; 5 — узел блуждающего нерва; б — синокаро-тидный нерв; 7 — наружная сонная артерия; 8—внутренняя сонная артерия
Влияние гипоксического фактора на дыхание выражается в его углублении. В то же время гиперкапнический фактор (избыток в крови углекислого газа) воздействует на дыхательный центр таким образом, что дыхание учащается.
Влияние состава крови на деятельность дыхательного центра было показано при помощи опыта с перекрестным кровообращением. У двух собак перерезают сонные артерии и соединяют их между собой таким образом, что кровь из туловища первой собаки течет в голову второй собаки, а кровь из туловища второй собаки — в голову первой собаки. Вторые сонные артерии зажимают (рис. 5.13). После установления перекрестного кровообращения между двумя собаками зажимают трахею первой собаки, т. е. производят ее удушение. При удушении первой собаки одышка наступает не у нее, а у второй собаки, голова которой снаб-
жается кровью первой собаки. Такое изменение дыхания объясняется тем, что при удушении первой собаки произошло изменение (повышение концентрации углекислоты) состава ее крови, а
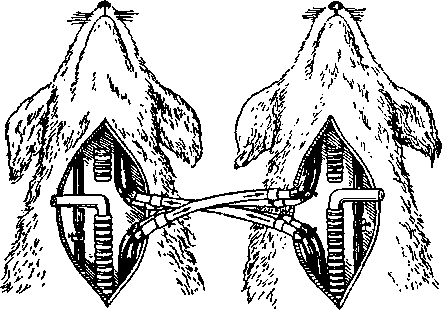
Рис.'5.13. Схема опыта с перекрестным кровообращением
так как эта кровь омывает головной мозг второй собаки, то она влияет на деятельность дыхатель-ного центра второй собаки, вызывая у нее одышку. Углекислота является специфическим возбудителем дыхания. Малейшие колебания ее концентрации в крови вызывают изменение деятельности дыхательного центра. Действие химических факторов в основном осуществляется через нервную систему рефлекторно. Это было показано в опытах, когда через сосуды различных органов, у которых с организмом была сохранена только нервная связь, пропускался раствор с повышенным содержанием углекислоты. Смывание органа подобным раствором вызывало учащение и углубление дыхания. Так же рефлекторно изменяется дыхание при недостатке кислорода в крови. Повышенная концентрация углекислоты в крови может оказать непосредственное действие на нейроны дыхательного центра. Если концентрация углекислоты в крови, омывающей дыхательный центр, повышается, то концентрация углекислоты в нейронах дыхательного центра также возрастает и дыхательный центр возбуждается, вследствие чего дыхание становится частым и глубоким. Так продолжается до тех пор, пока в результате усиленного дыхания концентрация углекислоты в крови вновь не снизится до нормального уровня. На понижение концентрации углекислоты в крови дыхательный центр отвечает понижением возбудимости, вплоть до полного прекращения своей деятельности на некоторое время, до установления нормального уровня углекислоты. Остановка дыхания, которая наступает в результате понижения концентрации углекислоты в крови, называется апноэ. Нарушение ритма дыхания — одышка и учащенное дыхание вследствие повышения концентрации углекислоты в крови — называется диспноэ. Явления апноэ и диспноэ можно изучить при помощи следующего простого эксперимента. Если в течение некоторого времени (1—2 минуты) часто и глубоко дышать, то после этого дыхание на некоторое время прекращается, т. е. наступает апноэ. Это связано с тем, что при частом и глубоком дыхании увеличивается вентиляция легких, а вместе с тем повышается количество выдыхаемого углекислого газа. Концентрация углекислоты в крови понижается, и в связи с этим возбудимость дыхательного центра тоже падает. Остановка дыхания продолжается до тех пор, пока в крови не накопится достаточное количество углекислоты. Наоборот, при задержке дыхания на 1—1 '/2 минуты наступает диспноэ — усиленное дыхание. Это объясняется тем, что при задержке дыхания концентрация углекислоты в крови повышается, вследствие чего увеличивается и возбудимость дыхательного центра. Усиленное дыхание продолжается до тех пор, пока концентрация углекислоты в крови не снизится.
Считается, что с повышением концентрации углекислоты в крови связан и первый вдох новорожденного. Как известно, в период внутриутробной жизни плод имеет с матерью общее кровообращение, осуществляемое через плаценту и сосуды пуповины. Все процессы обмена питательных веществ и газов совершаются через пуповину и плаценту. Плод получает из крови матери кислород и туда же отдает углекислый газ. Легкие же плода находятся в спавшемся состоянии. При рождении пуповину зажимают, перерезают, и связь с организмом матери прекращается. Прекращается приток кислорода и удаление углекислоты, но окислительные процессы в организме новорожденного продолжаются, и концентрация образовавшейся углекислоты при этом возрастает, так как она не удаляется из организма. В итоге концентрация углекислоты в крови достигает такой величины, что рефлекторно, а возможно, и непосредственным действием на нервные клетки дыхательного центра возбуждается его деятельность у новорожденного. Возбужденный центр дыхания, в свою очередь, посылает импульсы дыхательной мускулатуре, которая сокращается, грудная клетка расширяется и воздух устремляется в легкие, расправляя альвеолы. Так совершается первый вдох новорожденного ребенка.
Большой интерес представляют так называемые защитные рефлексы со слизистых оболочек дыхательных путей. Эти рефлексы называются защитными, потому что они либо препятствуют попаданию в дыхательные пути вредных веществ, либо способствуют удалению уже попавших г аздражающих веществ. Так, например, при вдыхании аммиака у человека наступает остановка дыхания после резкого выдоха и одновременно происходит рефлекторное сужение просветов брон-х jb и полное закрытие голосовой щели. Благодаря такой рефлекторной реакции, препятствующей попаданию в легкие вредных веществ, организм предохраняется от ядовитых летучих
веществ. При попадании пыли или слизи в гортань или бронхи рефлекторно возникает кашель, и чужеродное вещество с силой выталкивается из дыхательных путей. Перед кашлем человек производит вдох, и голосовая щель рефлекторно закрывается. Вслед за этим воздух судорожно выталкивается из легкого при широко раскрытой голосовой щели, что и обеспечивает звуковой эффект кашля. Вместе с вытолкнутым воздухом выводится из организма и раздражитель. При раздражении слизистой оболочки носа возникает чиханье. При чиханье, как и при кашле, человек производит глубокий вдох, при этом голосовая щель закрывается, мягкое небо поднимается и закрывает задний носовой ход. Затем происходит сильный выдох, воздух устремляется в задний носовой ход и оттуда через нос наружу с характерным звуком. При этом вещество, раздражавшее слизистую оболочку носа, удаляется.
Необходимо еще раз подчеркнуть, что в нормальном целом организме регуляция дыхания осуществляется при участии коры головного мозга. Человек может до определенного предела произвольно изменять частоту и глубину дыхания, что является результатом деятельности коры головного мозга. Влияние коры головного мозга на дыхание можно наглядно продемонстрировать при помощи следующего эксперимента. Если дать испытуемому вдыхать воздух с углекислым газом в концентрации 7—8% (углекислый газ вызывает учащение дыхания) и сочетать вдыхание углекислого газа со звуком метронома, то через несколько опытов один только звук метронома вызывает учащение дыхания. Это учащение наступает в результате образования условного рефлекса. Изменение дыхания, которое наступает до начала физической работы или спортивного упражнения только при одной команде, происходит также под влиянием коры головного мозга условнорефлекторно.
Разберем теперь, как происходит обмен газов между альвеолярным воздухом, т. е. воздухом находящимся в альвеолах, и кровью.
Человек дышит атмосферным воздухом, который имеет следующий состав: кислорода 20,94%, углекислого газа 0,03%, азота и инертных газов (аргон и др.) 79,03%. Состав же выдыхаемого воздуха следующий: кислорода 16,3% углекислого газа 4% и азота 79,7%. Надо учесть, что приведенные данные являются средними и колеблются довольно резко. Это непостоянство в составе выдыхаемого воздуха зависит от интенсивности обмена веществ, а также от частоты и глубины дыхания. Как видно из приведенных данных, а также из данных таблицы 5,1, где приведен состав вдыхаемого, выдыхаемого и альвеолярного воздуха, содержание азота в выдыхаемом и альвеолярном воздухе выше, чем в атмосферном. Эта разность не является следствием того, что азот вдыхается меньше, а выдыхается больше. Азот и инертные газы в газообмене не участвуют. Такая разница обусловлена тем, что объем вдыхаемого воздуха больше, чем выдыхаемого, так как кислорода потребляется больше, чем выделяется углекислого газа, и поэтому процентное содержание азота возрастает, в силу чего одно и то же количество его распределяется в меньшем объеме. Состав альвеолярного воздуха во многом отличается от атмосферного. В нем резко падает содержание кислорода и возрастает количество углекислого газа. Такое изменение состава объясняется тем, что именно здесь, в альвеолах, происходит обмен газов между воздухом и кровью, причем кровь поглощает кислород и отдает углекислый газ. Процентное содержание отдельных газов в альвеолярном воздухе следующее: кислорода 14,2—14,6%, углекислого газа 5,2—5,7% и — азота 79,7—80,6%.
Таблица 5.1