

3.2.3. Восприятие и распознавание речи
Слуховая система включает орган слуха и его связи с центральной нервной системой. С анатомической точки зрения ее можно подразделить на наружное, среднее и внутреннее ухо, слуховой нерв и центральные слуховые пути (Гельфанд, 1984) (Рис. 3.8.1, 3.8.2). Прием речевого сообщения начинается в наружном ухе, структуры которого (ушная раковина, наружный слуховой проход и барабанная перепонка) обеспечивают направленный прием звуковых колебаний из внешней среды. Наружный слуховой проход выполняет функцию резонатора, усиливающего звуковые частоты в диапазоне от 3,0 до 5,0 кГц приблизительно в 2—4 раза. В среднем ухе, представляющем костную полость, содержащую три слуховые косточки — молоточек, наковальню и стремечко, ограниченную с одной стороны барабанной перепонкой, с другой — костной полостью улитки, происходит увеличение уровня передаваемого давления приблизительно на 30 дБ. Главной функцией среднего уха является выравнивание рассогласования в акустическом сопротивлении воздуха и жидкости улитки.
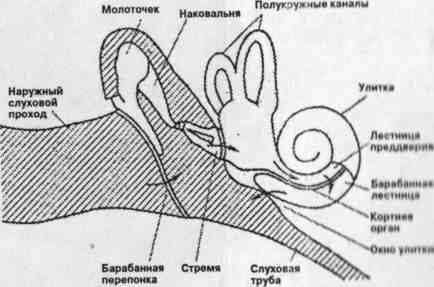
Рис. 3.8.1.
Необходимый баланс давления (давление воздуха во внутреннем ухе должно быть равно внешнему атмосферному давлению) поддерживается посредством евстахиевой трубы соединяющей среднего уха с носоглоткой. Внутреннее ухо представляет собой сложно организованный лабиринт каналов, расположенных в височной кости и заполненных жидкостью. Костная структура имеющая форму спирали, называется улиткой. Две мембраны проходят вдоль всей длины улитки от ее базилярной (основной) части до апикального конца. Соответственно они называются базилярной (основной) и Рейснерозой мембранами. Мембраны делят полость улитки на три канала: вестибулярный, тимпанальный (которые заполнены перилимфой) и средний (заполненный эндолимфой). Каналы соединяются у вершины улитки через небольшое отверстие — геликотрему. В среднем канале находится рецепторный аппарат слуха — кортиев орган. Реакция базилярной мембраны на импульс увеличения давления описывается как «бегущая волна» (Шупляков, 1990). Высокие частоты приводят к максимальному смещению мембраны у ее основания, понижение частот — к локализации максимального смещения к ее вершине. Верхняя половина мембраны реагирует на колебания с частотами от 0 до 2000 Гц, остальная часть — на все другие частоты. Наличие «точек» максимального смещения на каждую частоту лежит в основе «теории места». На колебательный процесс, возникающий на мембране, влияет интенсивность звука: чем сильнее звук, тем больше размах колебаний всех участков мембраны. Таким образом, связь местонахождения максимального возбуждения базилярной мембраны с частотой звука свидетельствует о частотно-избирательной, или фильтрующей функции этого органа слуха. Во внутреннем ухе осуществляется процесс перекодирования частоты и интенсивности звукового сигнала в активность, локализованную на определенном участке базилярной мембраны. Об особенностях частотного и временного анализа можно прочитать в руководстве по слуху (Шупляков, 1990). Однако следует иметь в виду, что частотно-избирательные колебания базилярной мембраны дают только грубую информацию о спектральных различиях между звуками. В то же время в целом внутреннее ухо работает как фильтр, пропускающий преимущественно экологически значимые частотные диапазоны, т.е. реализуется принцип согласования приема (Куликов, 1986).
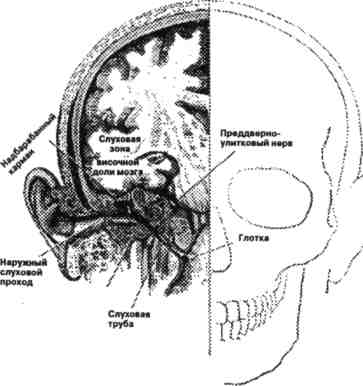
Рис. 3.8.2. Расположение структур уха по отношению к голове. (Источник перепечатки: Гельфанд С, 1984. С.12.)
Передача информации о физических параметрах звуковых колебаний в вышележащие отделы слуховой системы осуществляется посредством кортиева органа и связанных с ним волокон слухового нерва. Кортиев орган представляет собой сложную систему волосковых клеток-рецепторов, распределенных вдоль базилярной мембраны в два слоя (наружный и внутренний), выполняющих функцию электромеханических датчиков. Возбуждение волосковых рецепторов имеет форму электрической активности, которая передается окончаниям и волокнами слухового нерва, вызывая в нервных волокнах генерацию электрических импульсов. Афферентные волокна слухового нерва передают информацию на следующие уровни слуховой системы. Первые слуховые нейроны соединены с волосковыми клетками, при этом около 95% нейронов контактируют по принципу «один нейрон — одна (внутренняя) волосковая клетка, одна клетка — 10—12 нейронов». Такая организация обеспечивает связь «частота звукового сигнала—место на мембране—место в слуховом нерве» и дублирование информации. Около 5% нейронов соединены с 10—12 внешними волосковыми клетками. Эти нейроны имеют более низкие пороги срабатывания. Таким образом, на уровне первичных нейронов существует дде системы иннервации. Вследствие непосредственной связи между нейронами и определенными «точками» базилярной мембраны слуховой нерв можно рассматривать как ее проекцию, а совокупность нервных волокон, организованных по принципу «частота—место», как набор электрических фильтров-резонаторов, настроенных на определенную частоту. Порог чувствительности нейрона к тону данной частоты определяется минимальным уровнем интенсивности тона, при котором в нервном волокне возникает ответный импульс. Частота тона, соответствующая наименьшему порогу чувствительности, называется характеристической частотой нейрона (Чистович и др., 1976). Нейроны чувствительны к частотам, близким к их характеристическим частотам, и практически не реагируют на сигналы с частотами, большими характеристической частоты. Максимальную чувствительность к сигналу имеют нейроны с частотами в области речевых частотных составляющих— 1000—4000 Гц. Минимальные пороги чувствительности остальных частот выше в области низких частот.
Частотная полоса возбуждения нейронного фильтра зависит от интенсивности сигнала; при больших интенсивностях ширина полосы увеличивается. Для очень громких звуков это приводит к ухудшению частотной избирательности слуха.
Частота сигнала в значительной области звукового диапазона кодируется в слуховой системе местом активизированного нейрона или группы нейронов и периодом суммарного потока нейронной активности в возбужденной частотной полосе. Следовательно, в слуховом нерве происходит процесс пространственно-временного преобразования звуковых колебаний в нейронное возбуждение, передаваемое частотной им пульсацией.
Важнейшие свойства человеческого слуха, к которым относится оценка громкости, а также абсолютная и дифференциальная чувствительность к звуковым сигналам разной интенсивности и частоты, определяются вышеописанными реакциями базилярной мембраны и волокон слухового нерва. Под абсолютной чувствительностью подразумевается способность человека слышать разные звуки минимальной интенсивности, под дифференциальной — различать звуки с различной частотой и интенсивностью.
Обработка информации продолжается в лежащих выше центральных структурах слуховой системы, имеющих сложное нейроанатомическое строение. В этой многоуровневой организации на каждом уровне происходит обработка информации и последующая ее передача в вышележащие уровни (Рис. 3.9). Сохраняется настройка нейронов на определенные частоты и связь этих частот с, пространственным расположением нейронов и их группировок. Каждая группа нейронов обрабатывает информацию в ограниченной частотной полосе, а анализ спектральной информации на всем пути ее прохождения происходит в отдельных «сквозных» частотных каналах, в совокупности составляющий частотный диапазон человеческого уха. В первом приближении можно говорить о двух этапах обработки: в отдельных частотных каналах и путем выявления акустико-слуховых признаков речевого сигнала. В центральных отделах слуховой системы выделяют два типа нейронов: тонические и фазические (Чистович и др., 1976). Функция фазических нейронов заключается в выявлении изменяющихся акустических свойств сигнала.

Рис.3.9. Упрощенная схема слухового пути. (Источник: Квинке Р. Физиология слуха. В книге: Основы сенсорной физиологии. С.220.).
Эти нейроны характеризуются избирательным реагированием на увеличение и/или уменьшение энергии, изменение скорости. Фазические нейроны могут быть узкоспециализированными, откликающимися избирательно на изменение частоты тонального сигнала. В отклике тонических нейронов подчеркивается выраженность спектрального максимума в анализируемой полосе частот. Обнаружены также нейроны, реагирующие на периодичность гласного, что, по-видимому, играет роль в передаче информации о высоте голоса.
Процессы взаимодействия фазических и тонических нейронов при обработке речевого сообщения, а также вопрос об объединении акустических событий, достаточно сложны для интерпретации в данном изложении, так как требуют дополнительного представления сведений о нейрофизиологических особенностях нейронов центральных отделов слуховой системы.
В целом можно заключить (Кодзасов, Кривнова, 2001), что речевые сигналы обрабатываются с помощью общих слуховых механизмов. При обработке речевых сигналов нейронные структуры слуха с высокой надежностью выделяют те акустические признаки, которые необходимы и полезны для последующей фонологической интерпретации.
При оценке фонетической полезности акустических характеристик сигнала главным критерием является их роль в идентификации данного звука. Существует понятие акустических ключей, которые человек использует для опознания фонемного состава речевого сообщения. Акустическими ключами для идентификации гласных являются частотные и амплитудные значения двух первых формант. При этом, как правило, одна из формант является определяющей. Так, для гласных, различающихся по признаку ряда, существенной является частота F2, для различий по признаку подъема— частота F1. Ключи существуют и для идентификации согласных. Акустический анализ шумных согласных показывает, что от места образования зависят как физические свойства шумового отрезка согласного, так и характеристики переходов F2 на граничных участках соседних гласных (Кодзасов, Кривнова, 2001). Для восприятия согласных, различающихся местом образования, важны и форматные ключи, используемые для идентификации соседних гласных. Примером множественности акустических ключей является признак «глупость/звонкость» согласного. На восприятие данного противопоставления влияют, хотя и в разной степени, почти все акустические признаки, которые сопровождают его артикуляционную реализацию. К этим признакам (для сочетаний «взрывной согласный—гласный») относятся: длительность временного интервала между моментом взрыва и началом фонации; наличие начального перехода F1 в последующем гласном; длительность шумового участка и выраженность в нем формантных усилений; интенсивность шумового участка относительно интенсивности последующего гласного в области начальной метки последнего; длительность предшествующего гласного;
характер изменения частоты основного тона в области метки взрыва. Для фонологических признаков согласных характерна множественность ключей, что схематично можно представить как «один признак — много ключей». Однако вопрос о доминантном ключе и взаимоотношениях между разными ключами при идентификации одного признака в ряде случае сложен.
Еще одним свойством восприятия является категориальность. Под категориальностью восприятия понимают неспособность испытуемых различать стимулы, которые они относят к одному фонемному классу. Способностью к категориальному восприятию обладают новорожденные младенцы и некоторые животные (например — шиншиллы). Большинство акустических признаков, физические характеристики которых влияют на восприятие согласных, имеют категориальную природу.
В целом проблема восприятия может быть сформулирована в ряде нескольких общих вопросов (Бондарко, 1998). На основании каких акустических характеристик звука носитель языка относит его к той или иной фонемной категории? Каким образом из множества акустических признаков выделяются значимые для идентификации сообщения? Какой объем собственно акустической информации необходим для восприятия смысла передаваемого сообщения? Каким образом происходит восприятие звуков неродного языка? Многие другие вопросы имеют отношение к тому или иному из этих общих вопросов.
Представленные выше сведения о процессе речеобразования и восприятия звуков речи являются базовыми для рассмотрения различных речевых функций и становления речи в раннем онтогенезе. Использование онтогенетического и филогенетического подхода позволит пролить свет на еще многие вопросы, касающиеся речеобразования и речеспецифичности.
Основная литература
БатуевЛ.С. Высшие интегративные системы мозга. Л., 1981.
Бондарко Л.В. Фонетика современного русского языка. Учебное пособие. СПб.: Изд-во СПбГУ, 1998.
Вартанян И.А. Нейрофизиологические основы речевой деятельности. Механизмы деятельности мозга человека. 4.1. Нейрофизиология человека. Л.. Наука. 1988.
Гельфанд С.А. Слух. Введение в психологическую и физиологическую акустику. М.: Мир, 1984.
Деркач М.Ф., Гумецкий Р.Я., Губа Б.М., Чабан М.Е. Динамические спектры речевых сигналов. Львов, 1983.
ЗиндерЛ.Р. Общая фонетика. М.: Высшая школа, 1979.
Кодзасов СВ., Кривнова О.Ф. Общая фонетика. М., 2001.
Куликов Г.А. Нейрофизиологические основы обработки акустических сигналов. Акустика речи и слуха. Сборник трудов. 1986.
Шупляков B.C. Физиология периферического отдела слуховой системы. Слуховая система. Л.: Наука, 1990.
Фант Г. Акустическая теория речеобразования. М., 1964.
Фланаган Дж. Анализ, синтез и восприятие речи. М., 1968.
Чпопович Л.А., Венцов А.В., Гранстрем М.П.идр. Физиология речи. Восприятие речи человеком. Серия «Руководство по физиологии». М.: Наука, 1976.
Щерба Л.В. Русские гласные в качественном и количественном освещении /J Языковая система и речевая деятельность (1936) Л., 1974.
DaniloffR., Schuckers G., Feth L. The physiology of speech and hearing. 1980.
Kent R.D., Read Ch. The acoustic analysis of speech. 1992.
Lofqvist A., Gracco V. Tongue body kinematics in velar stop production: Influences of consonant voicing and vowel context. Phonetica. 51. 1994
Stevens K.N. Acoustical aspects of speech production. In Handbook of physiology. Washington. 1964.
Jakobson R., Halle M. Fundamentals of language. 1956.