

Расщепление липидов в желудочно—кишечном тракте
Расщепление липидов происходит в 12—перстной кишке, куда поступают липаза с соком поджелудочной железы и конъюгированные желчные кислоты в составе желчи. Эмульгирование жира — обязательное условие для переваривания, так как делает гидрофобный субстрат более доступным для действия гидролитических ферментов — липаз. Эмульгирование происходит при участии желчных кислот , которые из—за своей амфифильности, окружают каплю жира и снижают поверхностное натяжение, что приводит к дроблению капли.
Гидролиз жира осуществляется при участии панкреатической липазы , которая, сорбируясь на поверхности капель жира, расщепляет эфирные связи в триацилглицеринах (ТАГ). Жирные кислоты отщепляются прежде всего из a —положения. В результате образуется — диацилглицерин, затем b —моноацилглицерин, который является основным продуктом гидролиза:
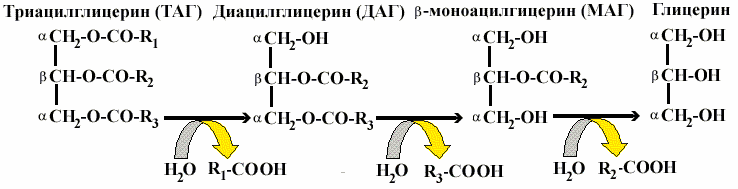
Всасывание происходит также при участии желчных кислот, которые образуют вместе с моноацилглицеринами, холестерином и жирными кислотами смешанные мицеллы — растворимые комплексы, обеспечивающие переход продуктов гидролиза в клетки слизистой кишечника. Желчные кислоты с током крови доставляются в печень, затем снова секретируются желчью в кишечник, то есть повторно используются, циркулируя по кругу: печень — кишечник — печень. Однако в течение суток примерно 0,3 г желчных кислот не всасываются, а выводятся с калом. Потери восполняются за счет синтеза в печени из холестерина. Нарушение желчеобразования или поступления желчи в кишечник приводит к нарушению расщепления жиров и их выделению в составе кала — стеаторрея .
Ресинтез
триацилглицеринов из продуктов
расщепления происходит в клетках
слизистой кишечника:

Транспорт ресинтезированного жира через лимфатическую систему и кровоток возможен только после включения его в состав липопротеинов.
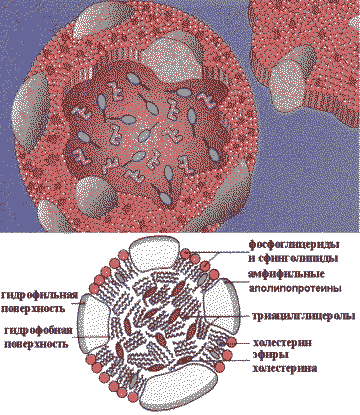
Строение липопротеинов
В кишечнике образуются два типа липопротеинов: хиломикроны — ХМ и в небольшом количестве липопротеины очень низкой плотности — ЛОНП. В составе хиломикронов экзогенные жиры доставляются в органы и ткани.
Липопротеинлипаза (ЛП—липаза) — фермент, обеспечивающий потребление экзогенных жиров тканями. ЛП—липаза, располагающаяся в эндотелии сосудов, взаимодействует с хиломикронами кровотока и гидролизует триацилглирины на глицерин и жирные кислоты, которые поступают в клетку. По мере извлечения ТАГ из хиломикронов последние превращаются в остаточные хиломикроны и затем поступают в печень. Потребность в жирах составляет 50—100 г. в сутки — в зависимости от характера питания и энергетических затрат. (14 вопрос)
Вопрос 14. Расщепление липидов в желудочно-кишечном тракте. Роль липолитических ферментов. Всасывание и транспорт липидов из кишечника в периферические ткани
РАСЩЕПЛЕНИЕ ПИЩЕВЫХ И ТКАНЕВЫХ ЛИПИДОВ
Пищевые липиды являются источниками высших жирных кислот, глицерола, аминоспиртов и некоторых других соединений, используемых организмом для синтеза свойственных для него структурных или резервных липидов. Свободные жирные кислоты, наряду с глицеролом и аминоспиртами образуются в организме также при расщеплении резервных или структурных липидов. Еще одним источником высших жирных кислот может служить их синтез из ацетил-СоА, который в свою очередь может быть промежуточным продуктом обмена углеводов или аминокислот (рис.21.1).
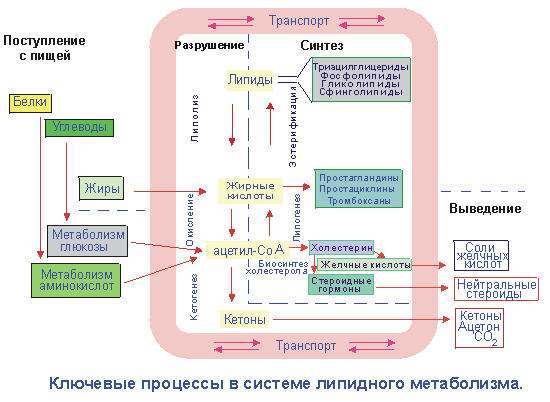
Рис. 21.1. Схема ключевых процессов липидного обмена
Одним из ключевых метаболитов липидного обмена является ацетил-СоА, поскольку, во-первых, именно через это соединение осуществляется окислительное расщепление высших жирных кислот; во-вторых, через ацетил-СоА атомы углерода жирных кислот могут быть использованы для пластических целей – для синтеза холестерола или полипреноидов; в третьих, через ацетил-СоА в гепатоцитах углеродные цепи жирных кислот преобразуются в кетоновые тела – гидрофильные «топливные» молекулы, легко транспортируемые в клетки различных органов и тканей; в четвертых, через ацетил-СоА осуществляются метаболические превращения углеродных скелетов аминокислот и моносахаридов в жирные кислоты (ЖК), используемые в дальнейшем для синтеза сложных липидов.
Соединения других классов – аминокислоты и моносахариды – в ходе своего метаболизма образуют промежуточные продукты, которые могут в дальнейшем использоваться в клетке как для синтеза высших жирных кислот, так и для образования других мономерных единиц, необходимых для синтеза сложных липидов: глицерола, этаноламина, холина, сфингозина и пр. Таким образом, обмен липидов оказывается тесно связанным с обменом соединений других классов, а метаболические пути обмена липидов различных классов являются частью метаболической сети, функционирующей в организме.
С пищей в организм человека ежедневно поступает от 80 до 150 г липидов животного и растительного происхождения. В составе липидов в организм поступают полиеновые жирных кислот, которые не синтезируются в организме. Кроме того, с липидами в организм поступают и жирорастворимые витамины – А, D, E и К. Основная масса липидов представлена жирами или триацилглицеролами. Они, наряду с глюкозой служат главными источниками энергии. На долю жиров при рациональном питании приходится не более 30% от общего числа калорий, поступающих с пищей. В пожилом возрасте, а также при малой физической нагрузке потребность в жирах снижается; в условиях физической работы – увеличивается.
Катаболизм липидов
В расщеплении простых и сложных липидов принимают участие липолитические ферменты, относящиеся к классу гидролаз, а сам процесс расщепления липидов носит название липолиза. В организме животных до 90% липидов, поступающих с пищей, приходится на долю жиров. Переваривание жиров происходит в тонком кишечнике. Предварительно нерастворимые в воде жиры эмульгируются. Эмульгирование происходит под действием солей желчных кислот, которые попадают с желчью в просвет 12-перстной кишки. Желчные кислоты действуют как детергенты, располагаясь на поверхности капель жира и снижая поверхностное натяжение. В результате крупные капли жира распадаются на множество мелких, т.е. происходит эмульгирование. Из крупной капли жира образуется 1012 мелких капель.
Гидролиз жиров осуществляется панкреатической липазой. Панкреатическая липаза выделяется в полость тонкой кишки из поджелудочной железы (ПЖЖ) в виде неактивной пролипазы. Превращение в активную липазу происходит при участии желчных кислот и еще одного белка панкреатического сока - колипазы. Этот фермент, также секретируемый в виде зимогена, активируется при гидролизе трипсином специфических пептидных связей. Активная колипаза образует с липазой комплекс в молярном отношении 1:1 за счет формирования двух ионных связей Lys-Glu и Asp-Arg.
Колипаза своим гидрофобным доменом связывается с эмульгированным жиром. Другая часть молекулы колипазы способствует формированию такой конформации панкреатической липазы, при которой активный центр фермента максимально приближен к молекуле жира, поэтому скорость гидролиза жира резко возрастает (рис.21.2).
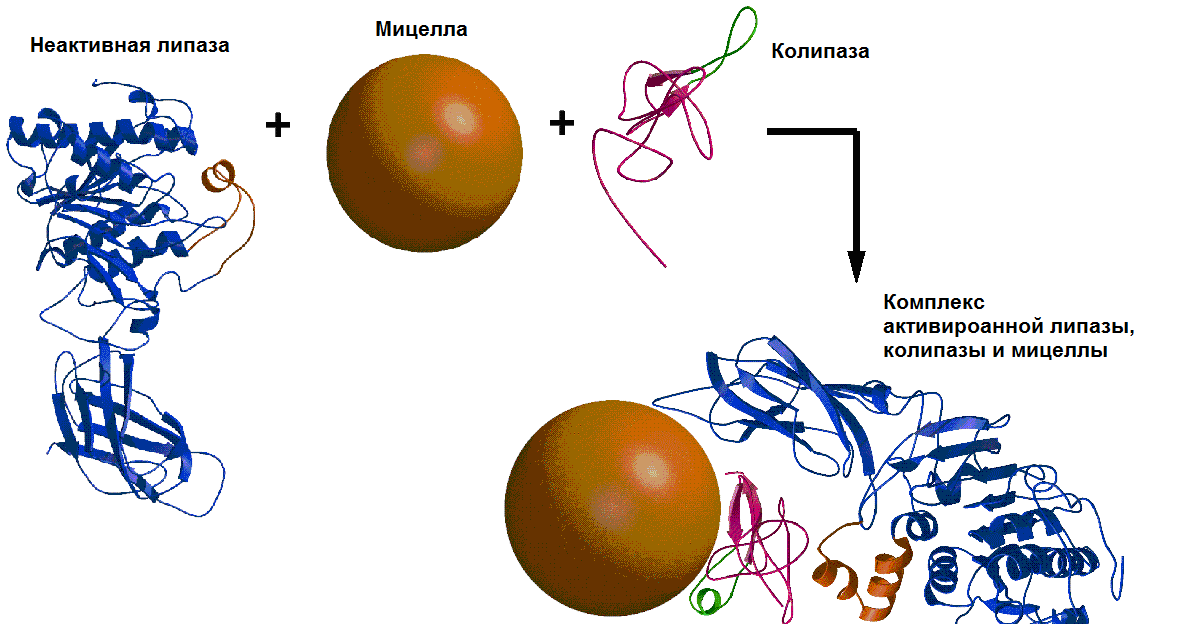
Рис.21.2. Взаимодействие неактивной панкреатической липазы, смешанной мицеллы и колипазы
Панкреатическая липаза гидролизует жиры преимущественно в 1 и 3 позициях (внешние сложноэфирные связи), поэтому основными продуктами гидролиза являются свободные ЖК и β-моноацилглицерол (2-моноацилглицерол, 2-МАГ). Молекулы 2-МАГ также обладают детергентными свойствами и способствуют эмульгированию жира.
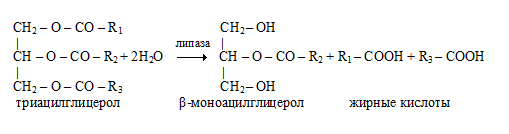
β-Моноацилглицеролы всасываются стенкой кишечника и либо участвуют в ресинтезе триацилглицеролов уже в кишечной стенке, либо распадаются до глицерола и высшей жирной кислоты под действием неспецифических эстераз.
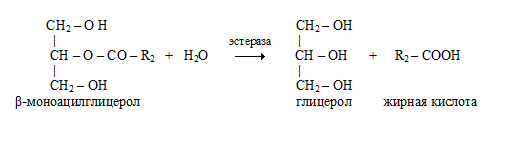
На скорость катализируемого липазой гидролиза триацилглицеролов не оказывает существенного влияния ни степень ненасыщенности жирной кислоты, ни длина ее цепи (от С12 до С18).
У растений в семенах и вегетативных органах присутствуют липазы, специфичность которых не выявлена. В дрожжевых грибках найдена липаза, способная отщеплять жирную кислоту как из α-, так и β-позиции.
Общепринято деление липаз на простые липазы, катализирующие гидролиз свободных триацилглицеролов, и липопротеинлипазы, гидролизирющие связанные с белками липиды.
Глицерофосфолипиды расщепляются под действием фосфолипаз. Существует четыре типа этих ферментов: фосфолипаза А1, А2, С и D. Фосфолипаза А1 отщепляет остаток жирной кислоты у С1 атома. Фосфолипаза А2 расщепляет β-сложноэфирную связь, фосфолипаза С отщепляет полярную головку вместе с остатком фосфорной кислоты, при этом продуктами гидролиза являются 1,2-диацилглицерол и фосфохолин. Фосфолипаза D, встречающаяся главным образом у растений, катализирует отщепление от фосфолипида полярной группы (азотистого основания) с образованием в качестве продукта фосфатидной кислоты (рис.21.3):
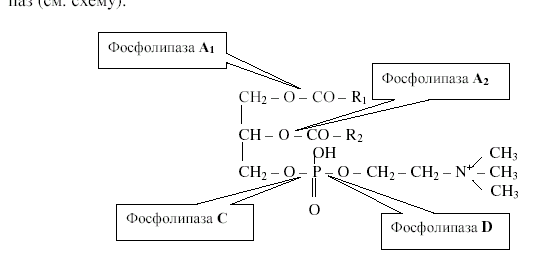
Рис.21.3. Расщепление сложноэфирных связей фосфолипазами
Фосфолипаза А2 (ФЛА2) гидролизует глицерофосфолипиды с образованием лизофосфолипидов. Лизофосфолипиды – эффективные эмульгаторы жира. Они, в свою очередь, под действием лизофосфолипазы, гидролизующей сложноэфирную связь у С1 атома, расщепляются на жирную кислоту и глицерофосфохолин, который хорошо растворяется в водной среде и всасывается из кишечника в кровь. Глицерофосфохолин также может расщепляться гидролазой до глицерол-3-фосфата и холина. ФЛА2 – неактивна, активируется путем частичного протеолиза, нуждается в Са2+.
Холестерол в пищевых продуктах содержится частично в свободном (неэстерифицированном) виде, частично в виде эфиров с жирными кислотами. Эфиры холестерола гидролизуются под действием особого фермента холестеролэстеразы, который синтезируется в ПЖЖ и секретируется в кишечник. Продуктами гидролиза являются свободный холестерол (ХС) и ЖК. Активность фермента проявляется в присутствии желчных кислот.
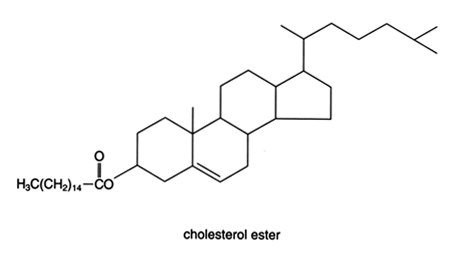
+ Н2О → СН3(СН2)14СООН
+
холестерол
Рис. 21.4. Гидролиз пальмитохолестерида холестеролэстеразой