

65. Кислотный метаболизм Толстянковых (сам-фотосинтез)
В сухих местах существуют растения-суккуленты, у которых устьица открыты ночью и закрыты днем для уменьшения транспирации. Поэтому эти растения поглощают углекислый газ ночью. Поглощение углекислого газа в темноте было обнаружено Н. Т. Соссюром (Швейцария) еще в 1804 г. Поглощаемый С02 используется, как и у С4-растений, для карбоксилирования ФЕП. ФЕП образуется ночью из крахмала в результате его гидролиза и включения образовавшегося глюкозо-6-фосфата в гликолиз. Катализирует реакцию карбоксилирования ФЕП-карбоксилаза. В результате образуется оксалоацетат и ортофосфат (рис. 4.18):
![]()
Оксалоацетат восстанавливается до малата при участии малат- дегидрогеназы:
![]()
Донором водорода является НАДН. Малат накапливается в вакуоли, поэтому в течение ночи клеточный сок становится все более кислым.
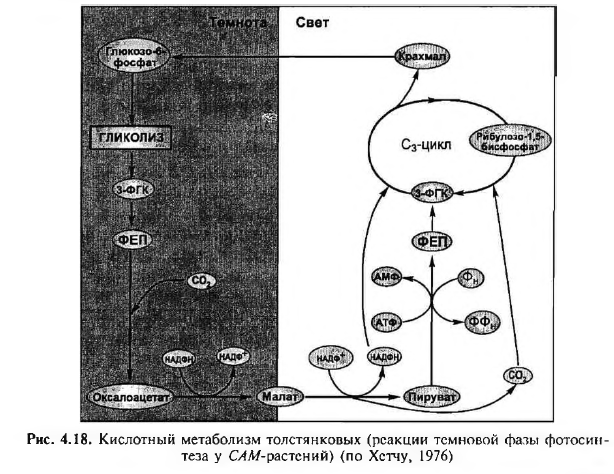
На следующий день утром малат выходит из вакуоли в цитозоль. Здесь происходит его окислительное декарбоксилирование при помощи НАДФ-малатдегидрогеназы (декарбоксилирующей), образуются пируват, С02, и восстанавливается НАДФ+:
![]()
Пируват фосфорилируется за счет АТФ с помощью пируват- фосфатдикиназы, и образуется ФЕП. С02 и НАДФН используются в С3-цикле.
Кроме того, на свету малат может вновь с помощью малатдегидрогеназы превратиться в оксалоацетат, который декарбоксили- руется под действием ФЕП-карбоксикиназы
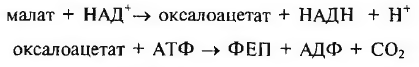
ФЕП-карбоксилаза, НАДФ-малатдегидрогеназа и ФЕП-карбоксикиназа находятся в цитозоле, поэтому катализируемые ими реакции происходят вне хлоропласта. По мере расходования кислот, накопленных ночью в вакуоли, величина pH клеточного сока вновь возрастает.
Образующийся после декарбоксилирования на свету малата или оксалоацетата ФЕП в результате ряда реакций превращается в ФГК или в глюкозо-6-фосфат. Днем малат ингибирует ФЕП-карбокси- лазу. ФГК может включиться в цикл Кальвина в хлоропласте (см. рис. 4.18), а глюкозо-6-фосфат — в первичный крахмал и в таком виде храниться, пока не наступит ночь. Таким образом, днем в хлоропластах этих растений идет обычный С3-цикл.
Так как этот тип фотосинтеза впервые был обнаружен М.Д. Хэтчем у растений из семейства толстянковых (Crassulaceae), то он получил название кислотного метаболизма толстянковых, или по-английски Crassulacean acid metabolism (CAM-фотосинтез), а имеющие его растения стали называть растениями CAM-типа, или САМ-растениями..
Для этого типа фотосинтеза характерны следующие особенности:
— темновая фаза фотосинтеза разделена во времени: С02 поглощается ночью, а восстанавливается днем. Для суккулентов это выгодно, так как позволяет днем закрывать устьица для уменьшения транспирации;
— из оксалоацетата — первичного продукта, как и у С4-растений, образуется малат;
— карбоксил ирование в тканях происходит дважды: ночью карбоксилируется ФЕП, днем — РуБФ.
Как и С4-ЦИКЛ, САМ-тип фотосинтеза является дополнительным, поставляющим С02 в Сз-цикл у растений, приспособившихся к жизни в условиях повышенных температур или недостатка влаги. Днем в жару С А Л/-растения запасают С02, образующийся в результате дыхания, а ночью, когда устьица открыты, они поглощают С02 из воздуха. Ночью С02 дыхания связывается ФЕП-карбоксилазой. Концентрация С02 в листьях САМ-растений днем колеблется от 0,5 до 2 %, а ночью падает ниже ее уровня в атмосфере. Повышенная концентрация двуокиси углерода способствует фотосинтезу. С4Л/-фотосинтез помогает уменьшать транспирацию и запасать воду, но продуктивность этих растений намного ниже, чем Сз-растений. Отношение транспирации к фотосинтезу для ОШ-растений составляет 50—100, для С4-растений — 250—300 и для С3-растений — 400—500 г транспирируемой воды/r поглощенного С02.
У большинства САМ-растений клетки мезофилла в несколько раз крупнее таких же клеток у С3- и С4-растений, так как у них очень большие вакуоли. Число устьиц у САМ-растений гораздо меньше, чем у С3- и С4-растений (табл. 4.4). В остальном анатомическое строение листьев у САМ-растений очень разнообразно.
В настоящее время этот тип фотосинтеза обнаружен у представителей 25 семейств. Среди изученных САМ-растений можно найти виды, у которых главной декарбоксилазой является НАДФ-малатдегидрогеназа (НАДФ-МДГ), и виды, у которых эту роль играет ФЕП-карбоксикиназа. Поэтому САМ-растения делят на 2 типа: НАДФ-МДГ-растения и ФЕП-КК-растения.
У некоторых растений этот цикл функционирует всегда, у других — только в неблагоприятных условиях. Переход к САМ-фотосинтезу происходит в условиях водного дефицита и высокой температуры днем и низкой ночью. Например, у ананаса наиболее интенсивная фиксация С02 в темноте наблюдалась при ночных температурах +5 — +10 °С. Переход к CW-фотосинтезу может вызвать короткий день (некоторые виды коланхое) или длинный (некоторые виды очитка). Существуют растения, например хрустальная травка, у которых можно вызвать переход от дневной фиксации С02 к ночной, поливая их соленой водой. Некоторые САМ-растения в условиях хорошего снабжения водой могут фотосинтезировать как С3-растения. Тогда у них устьица открываются днем и закрываются ночью. Иногда у САМ-растений С02 поглощается и днем, и ночью.