

62. Цикл Кребса. Механизмы регуляции цикла.
В начале цикла трикарбоновых кислот ацетил-кофермент А (ацетил-КоА) отдаёт свою ацетильную группу четырёхуглеродному соединению — оксалоацетату(щавелевоуксусной кислоте), при этом образуется шестиуглеродный цитрат (лимонная кислота). Ацетил-КоА является продуктом окисления таких соединений, как глюкоза, аминокислоты и жирные кислоты[6]. Цитрат затем изомеризуется в изоцитрат (изолимонную кислоту), который далее дегидрируется и декарбоксилируется до пятиуглеродной кислоты — α-кетоглутарата. α-Кетоглутарат вновь декарбоксилируется, превращаясь в четырёхуглеродный сукцинат (янтарная кислота). Сукцинат затем в три этапа ферментативно превращается в четырёхуглеродный оксалоацетат, который готов прореагировать с новой молекулой ацетил-КоА. В каждый оборот цикла одна ацетильная группа (то есть два атома углерода) приходит в цикл в виде ацетил-КоА, и два же атома углерода покидают цикл в виде двух молекул CO2; одна молекула оксалоацетата используется для образования цитрата, и одна же впоследствии регенерируется. Оксалоацетат не покидает цикл, и одна молекула оксалоацетата теоретически может связывать неограниченное количество ацетильных групп и, на самом деле, оксалоацетат присутствует в клетках в очень низких концентрациях. Четыре из восьми стадий цикла представляют собой окислительные процессы, выделяющаяся при этих процессах энергия окисления эффективно запасается в виде восстановленных коферментов НАДН и ФАДH2[5].
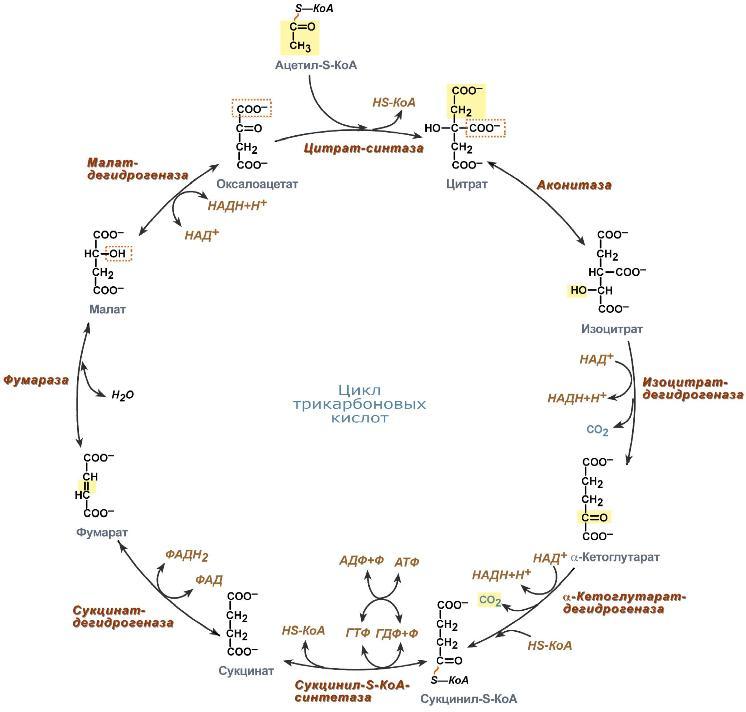
Регуляция цикла Кребса.
Дальнейшее использование образующегося из пирувата ацетил-СоА зависит от энергетического состояния клетки. При малой энергетической потребности клетки дыхательным контролем тормозится работа дыхательной цепи, а следовательно, реакций ЦТК и образования интермедиатов цикла, в том числе оксалоацетата, вовлекающего ацетил-СоА в цикл Кребса. Это приводит к большему использованию ацетил-СоА в синтетических процессах, которые также потребляют энергию.
Особенностью регуляции ЦТК является зависимость всех четырех дегидрогеназ цикла (изоцитратдегидрогеназы, а-кетоглутаратдегидрогеназы, сукцинатдегидрогеназы, малатдегидрогеназы) от отношения [NADH]/[NAD + ]. Активность цитратсинтазы тормозится высокой концентрацией АТР и собственным продуктом — цитратом. Изоцитратдегидрогеназа ингибируется NADH и активируется цитратом. а-Кето-глутаратдегидрогеназа подавляется продуктом реакции — сукцинил-СоА и активируется аденилатами. Окисление сукцината сукцинатдегидрогеназой тормозится оксалоацетатом и ускоряется АТР, ADP и восстановленным убихиноном (QH2). Наконец, малатдегидрогеназа ингибируется оксалоацетатом и у ряда объектов — высоким уровнем АТР. Однако степень участия величины энергетического заряда, или уровня адениновых нуклеотидов, в регуляции активности цикла Кребса у растений до конца не выяснена.
Регулирующую роль может играть также альтернативный путь транспорта электронов в растительных митохондриях. В условиях высокого содержания АТР, когда активность основной дыхательной цепи снижена, окисление субстратов через альтернативную оксидазу (без образования АТР) продолжается, что поддерживает на низком уровне отношение NADH/NAD+ и снижает уровень АТР. Все это позволяет циклу Кребса функционировать.