

Зависимость потока ионов от электрохимического градиента определяется уравнением Теорелла:

Уравнение Теорелла показывает, что два градиента являются движущей силой
потока ионов: градиент концентрации и градиент потенциала. Эти две величины: концентрация и потенциал, . являются главными составляющими энергии иона в среде, в том числе в липидной фазе мембраны, или, другими словами . составляющими электрохимического потенциала. Из сказанного ясно, что градиент электрохимического потенциала должен определять величину потока ионов. Его смысл достаточно ясен и интуитивно понятен: поток равен
произведению концентрации ионов, подвижности ионов и обратному градиенту
электрохимического потенциала.
где U - подвижность ионов, C - концентрация ионов, dμ/dx - электрохимический градиент.
Подставляя (6) в (7), можно получить уравнение Нернста-Планка с учётом двух градиентов, которые обуславливают диффузию ионов:
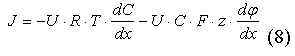
Активный транспорт ионов. Мембранный насос. Определение. Молекулярная конструкция натриево-калиевого насоса. Ионнообменный механизм транспорта ионов натрия, калия.
Активный транспорт — это перенос вещества из мест с меньшим значением электрохимического потенциала в места с его большим значением. Активный транспорт в мембране сопровождается ростом энергии Гиббса, он не может идти самопроизвольно, а только в сопряжении с процессом гидролиза аденозинтрифосфорной кислоты (АТФ), т. е. за счет затраты энергии
За счет активного транспорта в организме создаются градиенты концентраций, градиенты электрических потенциалов, градиенты давления и т.д.,
Существование активного транспорта веществ через биологические мембраны впервые было доказано в опытах Уссинга (1949 г.) на примере переноса ионов натрия через кожу лягушки
Согласно современным представлениям, в биологических мембранах имеются сложные белковые комплексы фермента АТФазы, работающие за счет свободной энергии гидролиза АТФ. Такие комплексы получили название ионных насосов
Мембранный насос-белок, содержащий активный центр, антиградиентная сила
Перенос ионов транспортными АТФазами происходит вследствие сопряжения процессов переноса с химическими реакциями, за счет энергии метаболизма клеток (см. термодинамическую модель переноса в разделе 23.9). На рис. 16.7 А показана наиболее распространенная модель K-Na-насоса. При работе К+-Ка+-АТФазы за счет энергии, освобождающейся при гидролизе каждой молекулы АТФ, в клетку переносятся два иона калия и одновременно из клетки выкачиваются три иона натрия. Таким образом, создается повышенная по сравнению с межклеточной средой концентрация в клетке ионов калия и пониженная натрия, что имеет огромное физиологическое значение.
В Са2+-АТФазе за счет энергии гидролиза АТФ переносятся два иона кальция, а в Н+-помпе — два протона на одну молекулу АТФ.
6. мембранный потенциал, определение, величина. Способы измерение МП.Условия и механизм возникновения МП. Роль пассивных и активных сил. Уравнение Нернста. Его природа. Стационарный МП, уравнение Гольдмана-Ходжкина.
мембранный потенциал - разность электрических потенциалов между наружной и внутренней поверхностями биологической мембраны, обусловленная неодинаковой концентрацией ионов, гл. обр. натрия, калия и хлора. На кривой потенциала действия различают восходящую и нисходящую фазы.
Уравнение Нернста — уравнение, связывающее окислительно-восстановительный потенциал системы с активностями веществ, входящих в электрохимическое уравнение, и стандартными потенциалами окислительно-восстановительных пар.

Измерение мембранного потенциала - получение количественных представлений о характеристиках мембранного потенциала.
Мембранный потенциал - условное название трансмембранной разности электрических потенциалов.
Все клеточные мембраны способны к электрической активности. Одним из свидетельств этому является наличие трансмембранной разности потенциалов (~30 ÷ 100 мв). Обычно эту разность потенциалов регистрируют с помощью двух электродов, размещенных на наружной и внутренней поверхностях плазмалеммы, присоединенных экранированными проводниками к электронному усилителю постоянного тока и, далее, к электронному осциллографу или иному регистрирующему устройству. Первый электрод, представляет собой микропипетку, заполненную электролитом. Тонкий конец пипетки имеет диаметр меньше 1 мкм. Этот электрод предназначен для введения в цитоплазму клетки и регистрации потенциала на внутренней поверхности плазмалеммы. Второй электрод используют для регистрации потенциала на наружной стороне плазмалеммы. Он может представлять собой серебряную пластинку. Его размещают во внеклеточной среде.
Когда электрическая цепь разомкнута или оба электрода находятся во внеклеточной среде, на экране осциллографа отмечается исходный нулевой уровень разности потенциалов. При проникновении первого электрода (микропипетки) внутрь клетки разность потенциалов скачком изменяется от исходного уровня до ~(-80) мв. Эту разность электрических потенциалов называют потенциалом покоя и обозначают: Ur , ПП (потенциал покоя), МПП (мембранный потенциал покоя), RMP (resting membrane potential).
Величина потенциала мембраны может изменяться от разных причин, главными из которых являются различные управляющие воздействия. Результатом этого может быть деполяризация (уменьшение поярности) или гиперполяризация (увеличение полярности). Управляемая полярность мембраны является основой возбуждения, возникновения потенциалов действия, проведения нервных импульсов, управления возбудимостью мембран, мышечного сокращения, секреции клеток желез, трансмембранного обмена веществ, восприятия, переработки информации и формирования управляющих воздействий и многих других неспецифических и специфических функций клеток, тканей, органов и систем органов.