

Лекция № 15. Тема «тканевое дыхание».
-
Состав и локализация дыхательной цепи:
а/ цитохромы, состав их молекул;
б/ роль убихинона в работе дыхательной цепи.
-
Состав и роль АТФ. (законспектировать из учебника).
-
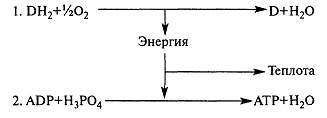
Окислительное фосфорилирование, его биологическая роль. Отличие от субстратного фосфорилирования.
-
Разобщение окислительного фосфорилирования.
-
Сущность хемиосмотической теории Митчелла.
-
Состав и локализация дыхательной цепи: а/ цитохромы, состав их молекул; б/ роль убихинона в работе дыхательной цепи.
Тканевое дыхание – это окисление органических веществ в организме кислородом с образованием воды и СО2.
Тканевое дыхание – это последовательность окислительно-восстановительных реакций, в результате которых происходит последовательный перенос водорода и электронов от субстрата к кислороду.
Тканевое дыхание включает:
-
Отнятие водорода от субстрата (дегидрогенирование);
-
Многоэтапный процесс переноса электронов на кислород;
Перенос электронов сопровождается уменьшением свободной энергии; часть этой энергии расходуется в виде тепла, а около 40 % используется на синтез АТФ. Тканевое дыхание и синтез АТФ энергетически сопряжены.
Ферменты отщепляющие водород от субстрата (дегидрогеназы), находятся в основном в матриксе митохондрий. Процесс тканевого дыхания протекает в митохондриях.
Всю систему реакций и ферментов тканевого дыхания называют дыхательной цепью.
Перенос электронов на кислород происходит при участии системы переносчиков, встроенных во внутреннюю мембрану митохондрий и образующих цепь переноса электронов (ЦПЭ).
В состав ЦПЭ входят 3 ферментных комплекса:
-
НАДН – дегидрогеназа (І);
-
QН2 – дегидрогеназа (ІІІ);
-
Цитохромоксидаза (ІV);
-
Низкомолекулярные переносчики: кофермент Q, цитохром с небольшой по размерам белок.
Направление переноса протонов и электронов определяет окислительно-восстановительный потенциал – редокс-потенциал. Все компоненты ЦПЭ расположены в митохондриях в порядке возрастания редокс-потенциала; самый высокий редокс - потенциал у кислорода. Это обеспечивает последовательное перемещение электронов от НАДН на кислород, при котором происходит выделение энергии на каждом этапе ЦПЭ.
Таким образом, работа дыхательной цепи сводится к транспорту электронов от субстрата тканевого дыхания к кислороду, а также протонов, поставляемых субстратом и матриксом митохондрий. При транспорте в связи с падением редокс-потенциала освобождается энергия и в итоге образуется вода.
Все участники дыхательной цепи (ЦПЭ) разделены на четыре окислительно-восстановительные системы (комплексы).
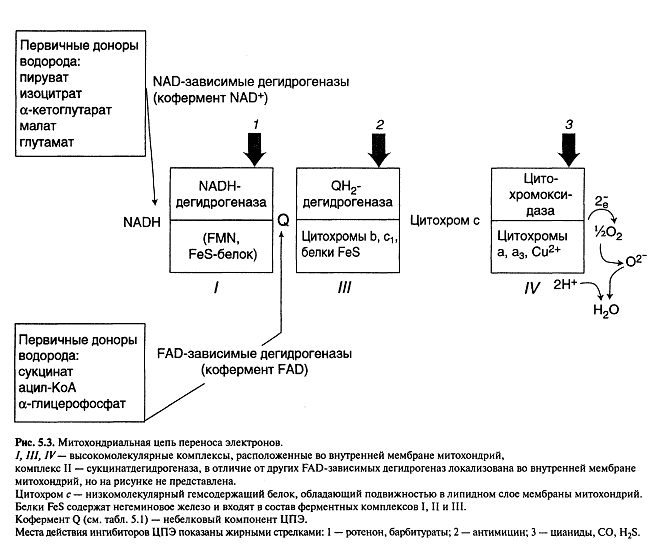
I комплекс дыхательной цепи: НАДН – убихинон – оксидоредуктаза. Это сложный полиферментный комплекс, обладающий большой молекулярной массой. Он содержит более 20 различных белков, коферментом которых является ФМН – флавинмононуклеотид, который способен восстанавливаться, присоединяя два атома водорода (т.е. 2 протона и 2 электрона), отдаваемых пиридиновыми дегидрогеназами (НАД – зависимая дегидрогеназа). ФАД + 2Н ↔ ФАДН2.
Кроме ФМН-зависимых ферментов, в состав I комплекса входят пять железосерных белков, которые осуществляют разделение потока протонов и электронов. При этом электроны от ФМН · Н2 переносятся к внутренней поверхности внутренней мембраны митохондрий (обращенной к матриксу), а протоны – к внешней поверхности внутренней мембраны, здесь они выделяются в межмембранное пространство.
Субстраты (SН2) проникают в матрикс митохондрии и подвергаются воздействию НАД+-зависимых дегидрогеназ:
SН2 + НАД+-ДГ-аза → S + НАДН-ДГ-аза + Н+.
Символом SН2 обозначаются такие субстраты, как пировиноградная кислота, изоцитрат, малат, оксипроизводные жирных кислот, глутаминовая кислота и другие аминокислоты.
Пиридиновые дегидрогеназы способны отнимать протоны и электроны у субстратов реакций, протекающих в матриксе, и передавать их первому комплексу дыхательной цепи (см. схему выше). Пиридиновые дегидрогеназы включают кофермент НАД+, в который входит витамин РР (производное пиридина). Окисленная форма (НАД+) называется пиридиниевым катионом.
В I комплексе дыхательной цепи непосредственно АТФ не образуется, а высвобождающаяся энергия аккумулируется или рассеивается в виде тепла.
II комплекс дыхательной цепи: сукцинат – убихинон – оксидоредуктаза.
Этот комплекс отличается меньшей молекулярной массой, содержит железосерные белки. С этим комплексом взаимодействует сукцинат, поступающий из матрикса митохондрий, а также жирные кислоты, которые находятся в матриксе. Коферментом комплекса является ФАД – флавинадениндинуклеотид.
В результате включения водорода субстратов через ФАД-зависимые дегидрогеназы (II комплекс) энергия в основном рассеивается в виде тепла, так как падение редокс-потенциала на этом участке дыхательной цепи незначительное (около 0,05 В) и этой энергии недостаточно для синтеза молекулы АТФ.
Убихинон (коэнзим Q) – посредник в передаче водородов. Это небольшая молекула (производное бензохинона) с длинной боковой цепью, способная свободно перемещаться как вдоль, так и поперек мембраны. Перемещаясь, молекулы убихинона захватывают протоны и электроны от комплексов I и II дыхательной цепи, а также протоны из матрикса. При этом убихинон восстанавливается.
КоQ + 2Н+ + 2е ↔ КоQН2
Восстановленная форма (убихинол, КоQН2) в свою очередь передает 2 электрона III комплексу дыхательной цепи (возможно, с участием какого-то индивидуального переносчика), а протоны при этом высвобождаются в межмембранное пространство.
III комплекс дыхательной цепи: убихинол – цитохром-с – оксидоредуктаза.
IV комплекс дыхательной цепи: цитохром-с – оксидаза.
В состав III и IV комплексов входят сложные белки – цитохромы. Цитохромы b и с1 объединены в III комплекс дыхательной цепи. Цитохромы а и а3 образуют IV комплекс дыхательной цепи.
Работа III комплекса заключается в транспорте электронов от убихинола на цитохрос с. Ферменты III комплекса способны захватывать из матрикса протоны и переносить их в межмембранное пространство. При этом существенно падает ОВ потенциал (от –0,04 В цитохрома «b» до +0,25 В цитохрома «с»), а высвободившейся энергии достаточно для синтеза одной молекулы АТФ.
От III комплекса электроны переносятся на IV при помощи очень подвижного фермента цитохромоксидазы (цитохром «с»). В своем составе этот фермент содержит 104 аминокислоты и одну гемподобную структуру. Такая молекула способна активно перемещаться, совершая челночные движения вдоль внешней поверхности мембраны от III к IV комплексу. Цитохром «с» при этом переносит только электроны, попеременно восстанавливаясь и окисляясь. В отличие от других цитохромов цитохромы а и а3 содержат, помимо железа, также медь, которая меняет свою степень окисления. При этом электроны от цитохрома «с» последовательно переносятся на цитохром «а», а затем на цитохром «а3».
Конечным акцептором электронов является молекулярный кислород воздуха. Восстановление кислорода происходит на цитохроме «а3», обращенном к матриксу:
2е + ½ О2 + 2Н+ → Н2О или
4е + О2 + 4Н+ → 2Н2О.
Ионы Н+ для образования молекул воды берутся из матрикса митохондрии. Редокс-потенциал IV комплекса велик (+0,57 В), его хватит на образование 2-х молекул АТФ. Важнейшей функцией IV комплекса является также активный транспорт протонов, в связи с чем этот дыхательный комплекс ферментов получил название «протонного насоса». Протоны транспортируются в межмембранное пространство митохондрий.