

Решение типовых задач
Задача 1. У бактерий Escherichia coli было получено 5 мутантов, затрагивающих разные этапы одного и того же биосинтетического пути (схема пути приведена на рис. 4), и изучена их способность к росту на среде, содержащей соединения (I–V), являющиеся предшественниками конечного продукта VI (табл. 3). Вам необходимо определить правильную очередность синтеза соединений I-VI в рассматриваемом биосинтетическом пути, а также идентифицировать мутанты, т.е. определить этап, где произошла мутация у каждого из них.
Таблица 3
Характеристика мутантов Escherichia coli
|
Мутанты |
Соединения |
|||||
|
I |
II |
III |
IV |
V |
VI |
|
|
1 |
- |
- |
- |
+ |
- |
+ |
|
2 |
- |
+ |
- |
+ |
- |
+ |
|
3 |
- |
- |
- |
- |
- |
+ |
|
4 |
- |
+ |
+ |
+ |
- |
+ |
|
5 |
+ |
+ |
+ |
+ |
- |
+ |
«+» – есть рост, «-» – нет роста.
Решение:
При решении задач подобного типа следует помнить, что если исследуемый ауксотрофный мутант растет на среде только в случае добавки необходимого метаболита, то у него блокирован какой-то этап биосинтетического пути на этапе, предшествующем этому метаболиту, то есть метаболит занимает в цепи место после мутационного блока. Другие более ранние предшественники, находящиеся в биосинтетическом пути перед блоком, не могут обеспечить рост мутанта. С другой стороны, если мутант накапливает определенный метаболит, то у него блокирован этап пути биосинтеза сразу же после накапливаемого метаболита. Например, в случае соединения V мы видим, что оно не обеспечивает рост ни одного из мутантов. Следовательно, в рассматриваемом биосинтетическом пути соединение V является самым первым предшественником. Затем следует соединение I, которое обеспечивает рост только одного мутанта (а именно, мутанта № 5), значит, в биосинтетическом пути оно располагается следующим. Затем следует соединение III и т.д. Следовательно, очередность синтеза соединений I-VI должна быть следующей: VIIIIIIIVVI.
Подобным образом рассуждаем и в случае идентификации мутантов. Поскольку мутант 5 не растет на соединении V, значит, мутация у него произошла на участке VI. Мутант 4 не растет уже на двух соединениях – V и I, следовательно, мутация у него произошла на следующем участке между IIII. Мутант 2 не может расти на соединениях V, I и III, следовательно, мутация у него произошла на участке IIIII. Рассуждая подобным образом и далее, определяем местоположение всех остальных мутаций.
Ответ.
Мутанты: 5 4 2 1 3
П



 уть:
V I
III
II IV
VI.
уть:
V I
III
II IV
VI.
Такой принцип идентификации мутантов применим для любого биосинтетического пути, и является универсальным как для прокариотических, так и эукариотических организмов.
З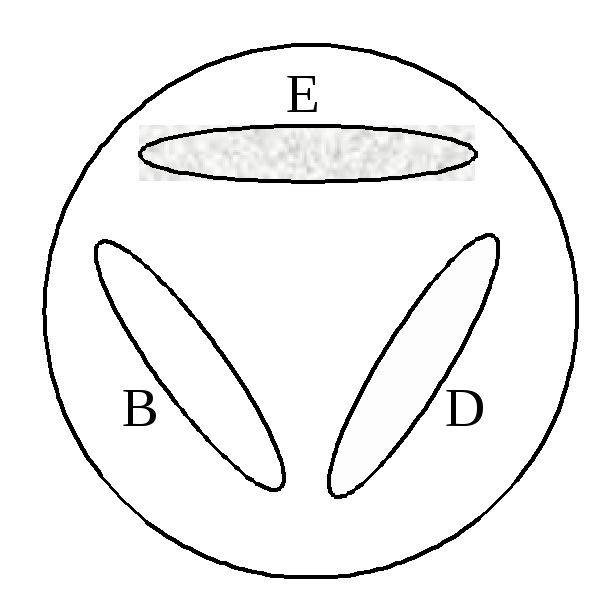 адача
2. Три
независимых триптофан-зависимых мутанта
дрожжей, обозначенных trp
B,
trp
D
и trp
E,
соответственно, были ресуспендированы
в физиологическом растворе и нанесены
на агаризованную среду без триптофана
в виде полосок, таким образом, что
отдельные штрихи не соприкасались между
собой, но могли использовать для своего
роста метаболиты других мутантов (рис.
5).
После выращивания мутантов в течение
2-3 суток был зарегистрирован рост
дрожжевых клеток, как указано на рисунке.
Проанализируйте результаты эксперимента
и определите, какова очередность действия
генов триптофанового пути у дрожжей?
адача
2. Три
независимых триптофан-зависимых мутанта
дрожжей, обозначенных trp
B,
trp
D
и trp
E,
соответственно, были ресуспендированы
в физиологическом растворе и нанесены
на агаризованную среду без триптофана
в виде полосок, таким образом, что
отдельные штрихи не соприкасались между
собой, но могли использовать для своего
роста метаболиты других мутантов (рис.
5).
После выращивания мутантов в течение
2-3 суток был зарегистрирован рост
дрожжевых клеток, как указано на рисунке.
Проанализируйте результаты эксперимента
и определите, какова очередность действия
генов триптофанового пути у дрожжей?
Рис. 3. Характер
роста мутантных
дрожжей (trp
B, trp D
и trp E)
на среде без триптофана.
Учитывая, что наличие роста на конкретном соединении говорит о том, что блок биосинтетического пути находится на более раннем его этапе, производим ана-
лиз картины роста мутантов дрожжей. Мы видим, что мутант trp E растет хорошо, и, следовательно, способен использовать для роста метаболиты мутантов trp D и trp B.
Значит в пути синтеза триптофана, фермент, кодируемый геном trp E, действует первым. Рост мутанта trp D слабый и неравномерный, этот говорит о том, что только один из двух мутантов обеспечивает его необходимым фактором роста. Поскольку мутант trp E имеет самый «ранний» блок триптофанового пути и не способен синтезировать метаболиты для «подпитки» других мутантов, следовательно, только клетки мутанта trp B синтезируют нужное для его роста соединение. Действительно, pост клеток мутантa trp D является более обильным в участке, приближенном к месту посева мутанта trp B. Значить фермент, кодируемый геном trp D, работает следующим.
Ответ. Последовательность генов в биосинтетическом пути:
trp E→ trp D → trp B.
Задача 3. В таблице приведены результаты теста на комплементарность для десяти точковых мутаций. «+» – комплементация мутации; «-» – отсутствие комплементации. По результатам, приведенным в таблице, определите группы комплементации.
|
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
1 |
- |
+ |
+ |
+ |
+ |
- |
- |
+ |
+ |
+ |
|
2 |
|
- |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
- |
|
3 |
|
|
- |
- |
- |
+ |
+ |
- |
- |
+ |
|
4 |
|
|
|
- |
- |
+ |
+ |
- |
- |
+ |
|
5 |
|
|
|
|
- |
+ |
+ |
- |
- |
+ |
|
6 |
|
|
|
|
|
- |
- |
+ |
+ |
+ |
|
7 |
|
|
|
|
|
|
- |
+ |
+ |
+ |
|
8 |
|
|
|
|
|
|
|
- |
- |
+ |
|
9 |
|
|
|
|
|
|
|
|
- |
+ |
|
10 |
|
|
|
|
|
|
|
|
|
- |
Решение:
1). Анализируем первую горизонтальную строку. Мы видим, что при скрещивании первого мутанта с шестым и седьмым мутантом не происходит комплементации мутации (знак «-»), следовательно, эти мутации затрагивают одну область гена, поэтому мы можем отнести эти мутации к одной группе комплементации. При скрещивании первого мутанта со вторым, третьим, четвертым, пятым, восьмым, девятым и десятым мутантом происходит комплементации мутации (знак «+»), следовательно, эти мутации затрагивают разные области гена, поэтому мы можем отнести эти мутации к другой группе комплементации.
2). Анализируем вторую горизонтальную строку. Мы видим, что при скрещивании второго мутанта с мутантом десять не происходит комплементации мутации (знак «-»), следовательно, эти мутации затрагивают одну область гена, поэтому мы можем отнести эти мутации к одной группе комплементации.
3). Анализируем третью горизонтальную строку. Мы видим, что при скрещивании третьего мутанта с мутантами четыре, пять, восемь и девять не происходит комплементации мутации (знак «-»), следовательно, эти мутации затрагивают одну область гена, поэтому мы можем отнести эти мутации к одной группе комплементации.
Ответ: Группа 1: мутанты 1, 6, 7.
Группа 2: мутанты 2, 10.
Группа 3: мутанты 3, 4, 5, 8, 9.
Задача 4. В результате эксперимента, получено пять делеционных варианта гена Х и пять точковых мутанта этого гена. Карта делеций представлена на рис.6. Пять точковых мутанта скрестили с делеционными вариантами. Результаты скрещивания представлены в таблице.
-
1

2
3
4
5
Рис.6. Делеционные варианты гена Х
-
Делеция
Точковый мутант
а
b
c
d
e
1
-
-
-
-
-
2
-
-
+
-
-
3
-
-
+
+
-
4
+
-
+
+
-
5
+
-
+
+
+
Решение:
Как указывалось выше, при скрещивании точкового и делеционного мутанта рекомбинанты появляются только тогда, когда делеция не затрагивает участок, в котором локализована точковая мутация. Делеции разделяют карту гена Х на пять областей (рис.6). Если при скрещивании точкового мутанта происходит рекомбинация только с некоторыми делециями, то эта точковая мутация локализована слева от самой протяженной делеции с которой произошла рекомбинация.
1. Анализируем результаты скрещивания мутанта а. Как видно из первого столбца таблицы при скрещивании точкового мутанта а с делеционными мутантами 4 и 5 происходит рекомбинация (знак «+»), а при скрещивании с делециями 1, 2 и 3.рекомбинация не происходит (знак «-»). Следовательно, данная точковая делеция локализована в области 3.
2. Анализируем результаты скрещивания мутанта b. Как видно из второго столбца таблицы при скрещивании точкового мутанта b с делеционными мутантами не происходит рекомбинация не с одной делецией (знак «-»). Следовательно, данная точковая делеция локализована в области 5.
3. Анализируем результаты скрещивания мутанта с. Как видно из третьего столбца таблицы при скрещивании точкового мутанта с с делеционными мутантами 2, 3, 4 и 5 происходит рекомбинация (знак «+»), а при скрещивании с делецией 1 рекомбинация не происходит (знак «-»).Следовательно, данная точковая делеция локализована в области 1.
4. Анализируем результаты скрещивания мутанта d. Как видно из таблицы при скрещивании точкового мутанта d с делеционными мутантами происходит рекомбинация мутации с делециями 3, 4 и 5 (знак «+»), но не рекомбинирует с делециями 1 и 2 (знак «-»). Следовательно, данная точковая делеция локализована в области 2.
5. Анализируем результаты скрещивания мутанта е. Как видно из таблицы мутант е рекомбинирует с делецией 5, но не рекомбинирует с делециями 1, 2, 3 и 4. Следовательно, данная точковая делеция локализована в области 4.
Ответ:
|
1 |
2 |
3 |
4 |
5 |
|
|
c |
d |
а |
e |
b |
|
Задача 5. В таблице представлены результаты попарных скрещиваний между пятью rII-делециями. «+» – комплементация мутации; «-» – отсутст-вие комплементации. Постройте карту делеций с указанием относительной длинны каждой из них. Какие из этих мутаций могли бы быть точковыми?
|
|
1 |
2 |
3 |
4 |
5 |
|
1 |
- |
- |
- |
- |
+ |
|
2 |
|
- |
- |
+ |
+ |
|
3 |
|
|
- |
+ |
+ |
|
4 |
|
|
|
- |
- |
|
5 |
|
|
|
|
- |
Решение:
1). Анализируем графы таблицы для определения самой протяженной делеции. Как видно из таблицы при скрещивании первого делеционного варианта с другими делециями комплементация происходит только при скрещивании с делецией 5 (знак «+»), следовательно, делеция 1 является самой протяженной.
2). Наносим на карту делецию которая комплементирует мутацию при скрещивании с самой протяженной. Это делеция 5. Отмечаем делецию 5 справа или слева от делеции 1.
3). При скрещивании делеции 2 с делецией 1 не происходит комплементации мутации (знак «-»), следовательно, она затрагивает делецию 1, но при скрещивании делеции 2 с делецией 5 происходит комплементация мутации (знак «+»), следовательно, она не затрагивает делецию делецию 5.
4). При скрещивании делеции 3 с делециями 1 и 2 не происходит комплементации мутации (знак «-»), следовательно, она затрагивает делецию 1 и 2, но при скрещивании делеции 3 с делецией 5 происходит комплементация мутации (знак «+»), следовательно, она не затрагивает делецию делецию 5.
5). При скрещивании делеции 4 с делецией 1 и 5 не происходит комплементации мутации (знак «-»), следовательно, она затрагивает эти делеции, но при скрещивании делеции 4 с делециями 2 и 3 происходит комплементация мутации (знак «+»), следовательно, она не затрагивает делеции 2 и 3.
Ответ. Карта rII-делециями делеций:
 1
1
2
3 5
4
Точковыми могли бы быть мутации 5, 3, 2.
Задача 6. У Pseudomonas putida получены ауксотрофные мутанты дефектные по различным этапам синтеза триптофана. Используя характеристику мутантов, приведенную в таблице, определите последовательность стадий биосинтеза триптофана у этих бактерий.
|
|
Рост на средах с предшественником |
||||
|
хоризмат |
антранилат |
КДРФ |
индол |
триптофан |
|
|
trpB |
- |
- |
- |
- |
+ |
|
trpE |
- |
+ |
+ |
+ |
+ |
|
trpC |
- |
- |
- |
+ |
+ |
|
trpA |
- |
- |
- |
+ |
+ |
|
trpD |
- |
- |
+ |
+ |
+ |
Решение:
Если мутант не растет на минимальной среде, а способен к росту только при внесении в среду определенного предшественника синтеза триптофана, то у него блокирован этап метаболической цепи перед биосинтезом этого метаболита.
-
Мутант trpE растет при добавлении в среду любого предшественника, кроме хоризмата, следовательно, у него блокирован самый ранний этап синтеза триптофана сразу после синтеза хоризмата.
-
Мутант trpD способен расти на средах со всеми промежуточными продуктами синтеза триптофана, кроме антранилата и КДРФ. Это означает, что мутационный блок располагается сразу после синтеза антранилата, который в цепи биосинтеза находится сразу после хоризмата.
-
Мутант trpC способен использовать для своего роста триптофан, и индол, но не антраниловую кислоту и КДРФ. Следовательно, мутация trpC прерывает цепь биосинтеза сразу после КДРФ, который располагается в цепи синтеза после антраниловой кислоты, но перед индолом и триптофаном.
-
Мутант trpA растет только на среде с триптофаном и индолом. Можно заключить, что цепь биосинтеза у него блокирована на этапе между индол-3-глицерофосфатом и индолом.
5. Мутант trpВ растет только на среде с триптофаном. Можно заключить, что цепь биосинтеза у него блокирована на этапе между индолом и триптофаном.
Таким образом, последовательность метаболитов в цепи биосинтеза триптофана следующая: хоризмовая кислота → антраниловая кислота → КДРФ →индол → индол-3-глицерофосфат → триптофан.