

Nb! в анаэробных условиях конечным акцептором водорода может быть ацетальдегид (спиртовое брожение глюкозы)
В анаэробных условиях глюкоза может превращаться в этанол. Ранее полагали, что образование этилового спирта – привилегия дрожжей и некоторых плесневых грибков. Однако уже доказано, что в тканях млекопитающих алкоголь также образуется. Он является нормальным метаболитом клеток. Тяга к алкоголю, по-видимому, возникает вследствие недостаточности ферментных систем, его производящих.
Реакции спиртового брожения глюкозы совпадают с реакциями гликолиза до стадии образования ПВК (рис.5.8).
Образование ацетальдегида из ПВК осуществляется путем прямого декарбоксилирования пирувата пируватдекарбоксилазой с участием ее кофермента – тиаминпирофосфата. Ацетальдегид превращается в этанол с помощью алкогольдегидрогеназы, коферментом которой является НАДН ·Н+. Уксусный альдегид – чрезвычайно токсичное соединение, его высокая концентрация в крови при потреблении спиртных напитков способна вызвать смертельный исход. Экзогенный этанол обезвреживается также алкогольдегидрогеназой (кофермент – НАД+), окисляясь в печени до ацетальдегида (реакция обратима) и далее с помощью альдегиддегидрогеназы – до уксусной кислоты. Активность альдегиддегидрогеназы (от ее зависит переносимость алкоголя) значительно варьирует у разных лиц и наций. Уксусная кислота, активируясь, превращается в ацетил-КоА, который “сгорает” в цикле Кребса с образованием энергии. В обезвреживании этанола принимают также участие микросомная система детоксикации гепатоцитов и фермент каталаза. Однако их роль менее значима.
Р ис.
5.8. Реакции образования этилового спирта
и его обмена
ис.
5.8. Реакции образования этилового спирта
и его обмена
Систематическое потребление алкоголя приводит к циррозу печени и увеличивает риск развития рака (причем не только печени), особенно на фоне хронического воздействия малых доз облучения.
Nb! Глюконеогенез – механизм синтеза глюкозы
Запасы гликогена в печени ограничены и после 12-18 часового голодания они исчезают полностью. Многие клетки нуждаются в постоянном обеспечении глюкозой (эритроциты, нейроны, мышечные клетки в анаэробных условиях). Глюконеогенез является тем метаболическим путем, который решает данную проблему. Глюконеогенез – это метаболический путь превращения неуглеводных соединений в глюкозу. Многие соединения могут участвовать в этом процессе. Это и молочная кислота, и ПВК, и аминокислоты, распадающихся до пирувата (аланин, цистеин, глицин, серин, треонин и др.), и глицерин, и пропиононил-КоА, и субстраты цикла Кребса (оксалацетат и др., рис. 5.8).
Глюконеогенез представляет собой модификацию таких процессов, как гликолиз и цикл Кребса. Большая часть реакций гликолиза обратима. Исключение составляют три реакции, которые катализируют гексокиназа, фосфофруктокиназа-1 и пируваткиназа и для преодоления этих реакций используются специальные ферменты, которые назвали ключевыми реакциями глюконеогенеза. Данные ферменты сосредоточены в печени и корковом веществе почек. В таблице 5.2. приводятся названия ферментов, катализирующих необратимые реакции гликолиза и соответствующих им ключевых ферментов глюконеоегенеза.
Таблица 5.2.Ключевые ферменты гликолиза и гликонеогенеза
-
Ферменты гликолиза
Ферменты глюконеогенеза
Гексокиназа
Глюкозо-6-фосфатаза
Фосфофруктокиназа-1 (ФФК-1)
Фруктозо-1,6-дифосфатаза
Пируваткиназа
Пируваткарбоксилаза
Фосфоенолпируваткарбоксикиназа
При совместной работе таких ферментов существует проблема т.н. “пустых” субстратных циклов. При условии катализа прямой и обратной реакции разными ферментами, продукт, получаемый в прямой реакции, становится субстратом другого фермента, который катализирует обратную реакцию, превращая продукт вновь в субстрат первого фермента. Возникает опасность “холостого” прокручивания субстратов реакции. Проблема решается организацией многоуровневой регуляции, включающей реципрокную аллостерическую регуляцию и ковалентную модификацию структуры ферментов.
Принято считать начальным
этапом глюконеогенеза реакции, идущие
в обход пируваткиназной реакции
гликолиза. Пируваткиназа – объект
влияния регуляторных систем(рис.5.9),
управляющих скоростью гликолиза, поэтому
в условиях благоприятствующих
глюконеогенезу (голодание и др.) активность
этого фермента следует затормозить.
Этому способствует повышение количества
аланина, который является аллостерическим
ингибитором пируваткиназы и усиление
секреции глюкагона. Последний стимулирует
образование цАМФ в гепатоцитах,
активирующей протеинкиназу А.
Фосфорилирование пируваткиназы под
влиянием протеинкиназы А вызывает
переход ее в неактивное состояние.
Торможение пируваткиназы благоприятствует
включению глюконеогенеза.
![]()
![]()
![]()
![]() .
.![]()
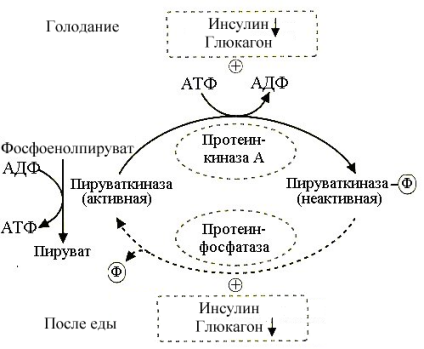
Рис.5.9. Регуляция активности пируваткиназы
Рис.5.10. Основные субстраты и ферменты глюконеогенеза:
1–лактатдегидрогеназа; 2– пируваткарбоксилаза; 3–малатдегидрогеназа; 4–фосфоенолпируват карбоксикиназа; 5–фруктозо-1,6-дифосфатаза; 6– глюкозо-6-фосфатаза; 7–глицеролкиназа; 8–-глицеролфосфатде гидрогеназа
Если превращение фосфоенолпирувата в ПВК, которое катализирует пируваткиназа, представляет одну химическую реакцию, то обратное превращение ПВК в фосфоенолпируват требует нескольких реакций. Первая реакция – это карбоксилирование пирувата. Реакция катализируется пируваткарбоксилазой и протекает с участием карбоксибиотина – активной форы СО2 в клетке. Продукт карбоксилирования – оксалоацетат занимает особое место в метаболизме митохондрий, где протекает данная реакция. Это важнейший субстрат цикла Кребса (см. ниже) и его выход из митохондрий затруднен. Для преодоления мембраны митохондрий оксалоацетат восстанавливается при помощи митохондриальной малатдегидрогеназы в легко приникающую через мебрану яблочную кислоту. Последняя, покинув митохондрии, в цитозоле окисляется вновь в оксалоацетат уже под влиянием цитозольной малатдегидрогеназы. Дальнейшее превращение оксалоацетата в ФЕПВК происходит в цитозоле клетки. Здесь при помощи фосфоенолпируваткарбоксикиназы окалоацетат декарбоксилируется с затратой энергии, высвобождаемой при гидролизе ГТФ и образуется ФЕПВК.
После образования ФЕПВК последующие реакции представляют обратимые реакции гликолиза. Из каждых двух образующихся 3-ФГА одна молекула при участии фосфотриозоизомеразы превращается в ФДА и обе триозы под влиянием альдолазы конденсируются в фруктозо-1,6-дифосфат. Некоторое количество ФДА образуется путем окисления глицеролфосфата, возникающего под влиянием глицеролкиназы из глицерола, поступающего в печень из жировой ткани. Это единственный субстрат из липидов, который участвует в глюконеогенезе. Превращение фруктозо-1,6-дифосфата во фруктозо-6-фосфат катализируется фруктозо-1,6-дифосфатазой-1. Затем вновь следует реакция, обратная гликолизу. Заключительная реакция глюконеогенеза катализируется ферментом глюкозо-6-фосфатазой, который катализирует гидролиз глюкозо-6-фосфата и образующаяся свободная глюкоза может выходить из клетки.
Суммарная реакция синтеза молекулы глюкозы:
2 ПВК + 4 АТФ + 2 ГТФ + 2НАДН + 2H+ + 6H2O → Глюкоза + 2НАД+ + 4АДФ+ 2 ГДФ + 6 Фн +6H+
Таким образом, синтез одной молекулы глюкозы “обходится” клетке затратой шести макроэргов. 2 молекулы АТФ расходуются для активирования СО2, 2 молекулы ГТФ используются в фосфоенолпируваткарбоксикиназной реакции и 2 молекулы АТФ – для образования 1,3-дифосфоглицериновой кислоты.
Глюконеогенез активируется в клетках печени во время голодания, после продолжительных физических упражнений, при употреблении пищи, богатой белками при низком содержании в ней углеводов и т.д.
Интенсивность процесса зависит от количества субстратов, и активности, и количества ключевых ферментов гликолиза и глюконеогенеза.
Основными поставщиками субстратов для печени являются мышцы, эритроциты, жировая ткань. У последней довольно ограниченные возможности, поскольку только глицерол может использоваться для синтеза глюкозы, а это только около 6% от веса капельки жира.
Лактат, образующийся в результате работы мышц в анаэробных условиях или поступающий из эритроцитов, более значимый источник глюкозы. Наиболее важными источниками являются гликогенные аминокислоты, которые могут поступать с пищей, богатой белками или из мышц в условиях голодания.
Рис. 5.11. Цикл Кори
Ч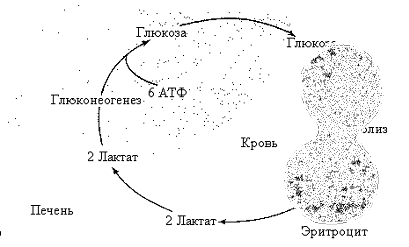 тобы
непрерывно снабжать глюкозой клетки,
для которых она является основным
источником энергии, но они не могут
окислить ее полностью в силу отсутствия
митохондрий (эритроциты) или из-за работы
в анаэробных условиях, между печенью и
этими клетками устанавливаются
циклические процессы по обмену
субстратами. Один из таких – цикл Кори:
образующаяся в мышцах (эритроцитах)
молочная кислота поступает в общий
кровоток, захватывается печенью и
используется ею в качестве субстрата
глюконеогенеза; синтезируемая при этом
глюкоза отдается в кровототок и
метаболизируется мышцами или эритроцитами
для получения энергии (рис. 5.11).
тобы
непрерывно снабжать глюкозой клетки,
для которых она является основным
источником энергии, но они не могут
окислить ее полностью в силу отсутствия
митохондрий (эритроциты) или из-за работы
в анаэробных условиях, между печенью и
этими клетками устанавливаются
циклические процессы по обмену
субстратами. Один из таких – цикл Кори:
образующаяся в мышцах (эритроцитах)
молочная кислота поступает в общий
кровоток, захватывается печенью и
используется ею в качестве субстрата
глюконеогенеза; синтезируемая при этом
глюкоза отдается в кровототок и
метаболизируется мышцами или эритроцитами
для получения энергии (рис. 5.11).
Р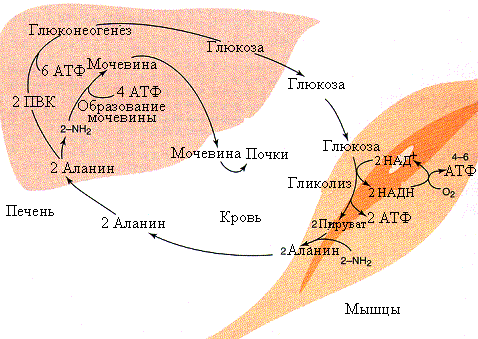 ис.5.12.Аланиновый
цикл
ис.5.12.Аланиновый
цикл
В отличие от цикла Кори, аланиновый цикл(рис.5.12) протекает при условии потребления периферическими тканями кислорода и требует митохондрий. При употреблении пищи богатой белами или при голодании происходит довольно активный обмен между печенью и мышцами аланином и глюкозой. Аланин из мышц передается клеткам печени, где он переаминируется и ПВК используется для синтеза глюкозы. По мере необходимости глюкоза поступает в мышцы и окисляется до ПВК, а затем, путем переаминирования, превращается в аланин который может вновь повторить этот цикл. Энергетически это более выгодный путь, чем цикл Кори.