

|
Билет 1 Термодинамическая система-совокупность макроскопических тел,которые могут взаимодействовать между собой и другими телами.Обмениваться с ними энергией и в-вом. ТДС состоит из большого числа молекул,атомов,структурных единиц,что его состояние можно хар-ть макроскопическими хар-ми:обьемом,давление,тем-ра.,плотность. концентрацией. Виды ТДС 1.Изолированные (нет обмена ни веществом ,ни энергией с окр.ср.) 2.Закрытые(нет обмена с в-вом с окр.средой) 3.Открытые( есть энерго и массообмен) Внутренняя энергия тел включает в себя: 1.Кинетическую энергию цепи движений части. 2.Потенциальную энергию заключ. в хим. связях. 3.Внутриядерную энергию.
В системах
хим.состав хоть не изменен, в процессе
энергетич.преобразований,можно
учитывать только тепловую энергию у
примеру удельный газ. U= Внутренняя энергия состоит из 2х частей: 1.Свободная энергия Gi-которую можно исп-ть для сов.работы.
2.Связанная энергия
Gc
связи-которую в данных условиях вообще
нельзя превратить в работу.U=Gi
+
Мерой связанной
энергии системы явл-ся энтропия
S= Энтропия S-величина связанной энергии происходящая на ед.абсолютной температуры. Те
процессы ,которые A= |
Билет №2 Первое начало термодинамики - представляет собой закон сохранения энергии. Он гласит, что энергия не может быть создана или уничтожена, она лишь переходит из одного вида в другой в различных физических процессах. Отсюда следует, что внутренняя энергия изолированной системы остается неизменной. В закрытых системах: изменение внутренней энергии; а)путем совершения работы(А); б)препятствует передачи тепла(Q). ∆U=Q-A Тепло, переданное системе, идет на VB и совершение работы Q=∆U+A +Q – тепло, поступающее в систему -Q – тепло, отдаваемое системой +A - работа совершаемая системой над телами -А - работа совершаемая телами над системой
Первое начало ТД
справедлив и для живых организмов, и
был сформулирован Р. Майср и Г. Геймгольд,
но записан в несколько другой форме
с учетом того, что единый источник
энергии для живых организмов является
энергия химических связей пищевых
продуктов.
Энергия пищи
тратиться на совершение работы,
изменение внутренней энергии и отдачи
тепла. Теплокровные имеют постоянную
температуру тела, и поэтому ∆U=0
,
Изотермический процесс(const= T) Физический смысл- все количество теплоты передаваемое системе, расходуется на совершение механической работы(расширение). Вся работа внешних сил выделяется в виде тепла(сжатие) Изохорический процесс(const=V) Физ. смысл-Все количество теплоты передаваемое системе расходуется (нагревание) на увеличение ∆V. Система уменьшает свою ∆V отдавая тепло(охлаждение) Изобарный процесс(const=p) Физ. смысл- Все количество теплоты передаваемое системе превышении совершаемой ею мех. работу. Количество теплоты отдаваемое системой превышает работу внешних сил Адиабатный процесс Физ. смысл- Совершение механической работы происходит путем уменьшения ∆V(расширение) V увеличение за счет работы внешних сил.
|
|
Билет 3 В живых организмах роль промежуточного звена между источником свободной энергии и работой в них выполняет не тепловая, а хим-кая энергия. В живых. Орг. работа не может совер-ся за тепловой энергии,тепло всегда означает тепловые потери при различных видах работы. Свободная энергия получаемая посредством пищи выделяется при биоокислении на мембранных митохондриях 30% рассеивается в виде тепла,ост.50% тратятся на синтез. Основные виды работ совершаемые в живых орг.: 1.Механическая работа. 2 Химический синтез. 3Поддержка физико-хим.градиента. 4.Перенос эл. эарядов.
|
Билет 4 Тепловой баланс организма. Вся тепловая энергия обр-ся в организме человека и уходит из него. Чтобы обеспечить постоянство t ры ,тело человека постоянно поддерживает равенство теплопродукции и теплообмена в соответствии с управлением теплового баланса.
M± Основные способы теплообмена орг-ма. В орг-ме любого живого сущ-ва непрерывно выделяется тепло. Это тепло должно отводиться в окр. среду, иначе орг. перегреется и погибнет, но однако слишком быстрая отдача тепла опасна и приводит в переохлаждению. Существуют два способа теплообмена орг-ма с окр.средой:
1.Теплопроводность-перенос
тепла за счет усиления молекулярного
движения в веществе Q=k* 2.Конвекция-перенос тепла связанный с движением газа или жидкости.
3.Излучение-
4.Испарение - Температурный гомеостаз Тем-ра тела человека и многих животных поддерживается постоянной с достаточно высокой точностью. Это св-во орг-ма называют температурный гомеостаз, которое обеспечивается выработкой в ходе эволюции системой терморегуляции. Способы терморегуляции. -Хим.терм-ция основана на изм-нии теплопродукции(скорости и хар-ра биоокисления, совершаемая мышечной работой -Дрожь -Физ.терморегуляция основана на изменении теплообмена.
|
|
Билет№5 Основный обмен. Понятия о методах измерения теплопродукции организма. Тепловая энергия, выделяемая организмами в окружающую среду в медицинской биофизике называют энерготратами организма. Энергозатраты сильно изменяются в зависимости от условий в которых пребывает человек. Энергозатраты бодрсвствующего организма(не во время сна) в условии исключении влияния внешних факторов называют основным обменом. Основной обмен зависит от возраста, чем старше человек, тем ниже значение ОО. Т.к. при старении митохондрии уменьшаются в количестве.
Расчет ОО=
Методы измерения теплопродукции -Прямая калориметрия основана на измерении тепла выделяемого организмом. В калориметре впервые был измерен ОО -Непрямая
калориметрия основана на исследовании
газообмена организма между
|
Билет№6 Понятие энтропии. Как мера связанной энергии. Вычисление через количество тепла переданное в процессе 2 начала ТД для изолированных систем.
Мерой связанной
энергии( Таким образом, энергия, в различных формах имеет разную практическую ценность. Чем меньше доля связанной энергии, мерой которого является энтропия, тем большую часть этой энергии можно преобразовать в работу. Следовательно, чем ниже энтропия системы, тем выше ценность свойственной ей внутренней энергии. При измерении энтропии рассчитывают:
а) при изотермическом
процессе ∆
б)в случайном
изменении температуры ∆ Второе начало ТД: в изолированной системе общее изменение энтропии всегда положительно. |
|
Билет №7 Вероятностный смысл энтропии. Флуктуация. Упорядоченность структур в свете 2 начала ТД. Состояние системы(макросостояние) характеризуется такими параметрами как T,P,V… Однако каждому макросостоянию системы, соответствует множество микросостояний, характеризуется различными положениями и скоростями микрочастиц образующих систему.
Степень упорядочности
системы характеризуется ее энтропией.
Поэтому между энтропией и вероятностью
состояния существует определенная
зависимость которую вывел Больцман
формулой: S=k*ln
Pтg
, где к-постоянная Больцмана k=R\
Pтg—термодинамическая вероятность – число микросостояний которыми может быть реализовано данное макросостояние
|
Билет 8
Все живые орг-мы
относятся к открытым системам, запас
свободной энергии открытой системе
в т.ч. и человека может как расти ,так
и убывать.Все будет зависеть от того,
что будет преобладать : приток свободной
энергии из окр.среды или убыль ∆
∆S=∆ Процессы внутри системы Процессы вне системы
Идут с изменением
(уменьш)свободной
получают ток
энергии за счет
процессов,поэтому∆
В открытой системе
изменение внутренней энергии за счет
пр.внутри системы всегда отрицательно
, а изменения энтропии за счет процессов
внутри системы положительно ∆
Продукция и поток энтропии Под продукцией энтропии понимается изменение энтропии вследствие процессов происх. Внутри нее.Поток энтропии выражает ту часть скорости изменения энтропии той же системы, которая обусловлена ее обменом энергией с окр.средой.
Скорости изм.
Энтропии
Стационарными состоянием системы называют состояние, в котором процессы в системе так сбалансированы, что основные величины,хар-щие систему,остаются постоянными. |
|
Билет№9 Структура. Физико-химические свойства биомембран. С точки зрения ТД живая клетка представляет собой открытую ТД систему. Биомембрана – это функционально активная белково-липидная структура, образующая границу раздела между клеткой и окружающей средой. Состоит из билипидного слоя, белков, полисахаридов гликокаликса. Модели:1)Жидкостно-мозаичная 2)Липопротеиновый коврик 3) «Сэндвич» Фосфолипиды: тело-глицерин, сфитозин:головки-азотистые основания(полярные) Хвосты- неполярные цепи нуклеиновых кислот.
|
Билет №10 Пассивный транспорт. 1)Перенос веществ по преобладающему физико-химическому градиенту 2)Без затраты энергии АТФ непосредственно на процесс переноса веществ через мембрану.
Понятие
физико-химического градиента- градиент
физической величины называют скорость
изменения этой величины в пространстве
grad
C=
Виды пассивного транспорта:1)Свободная диффузия липофильных веществ и через фосфолипидный бислой 2)Облегченная диффузия неэлектролитов 3)Электродиффузия, перенос ионов Диффузия- процесс переноса веществ из области с большой концетрацией в область с меньшей концетрацией за счет теплового движения молекул. Диффузия незаряженных частиц уменьшается в сторону этого градиента до тех пор, пока не состояния равновесия, пассивный транспорт, поскольку не требует затрат внешней энергии.
Характеристика
диффузии – поток вещества(φ)масса
переносимая через поверхность S
за единицу времени φ=
Отношения потока
вещества к площади плотности потока
j=
Уравнение диффузии
Фика j=-D
«=» - показывает
направление потока в сторону уменьшения
концетрации( т.е. против gradC)
D-коэффициент
диффузии D=
Для биомембран
существенное значение имеет коэффициент
распределения вещества между липидными
слоями и водой. Поэтому j= Посредством простой диффузии через фосфолипидный бислой проникают низкомолекулярные гидрофобные органические вещества(жирные кислоты)
|
 :
: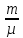 RT(i-число
степеней молек. µ-молекулярная масса.RT
RT(i-число
степеней молек. µ-молекулярная масса.RT
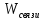 /T
/T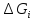 назыв-ся обратимой, т.к. пустив такой
процесс в обратном направлении и
затратив работу можно полностью
вернуть систему в исх.состояние,но
такие процессы в природе не сущ-ют.
Все реальные
процессы необратимые .Иными
словами,свободная энергия системы не
может быть преобразована в работу.
При таком преобраз. Часть
назыв-ся обратимой, т.к. пустив такой
процесс в обратном направлении и
затратив работу можно полностью
вернуть систему в исх.состояние,но
такие процессы в природе не сущ-ют.
Все реальные
процессы необратимые .Иными
словами,свободная энергия системы не
может быть преобразована в работу.
При таком преобраз. Часть
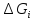 превращается в связанную,это является
диссипацией свободной энергии.
превращается в связанную,это является
диссипацией свободной энергии.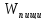 =A+Q+∆U
=A+Q+∆U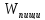 =A+Q
=A+Q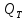 ±
± ±
±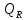 -
-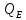 =0(М-теплопродукция,
Т-теплопроводность, С-конвекция
R-теплоизлучение,
Е-испарение.)(перенос тепловой энергии)
=0(М-теплопродукция,
Т-теплопроводность, С-конвекция
R-теплоизлучение,
Е-испарение.)(перенос тепловой энергии)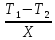 *S*t
*S*t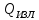 =Ӫ*(
=Ӫ*(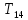 -
-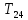 )
*S*t
)
*S*t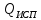 =l*m
(m-масса
исп.воды l-удельная
теплота испарения воды( 2.25*
=l*m
(m-масса
исп.воды l-удельная
теплота испарения воды( 2.25*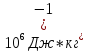 )
)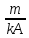 А-года. Коэффициент для мужчин:0,1015;
для женщин:0,1118
А-года. Коэффициент для мужчин:0,1015;
для женщин:0,1118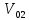 потребленного организмом и энерготратами
организма существует линейная
зависимость. Э\Т=20,2*
потребленного организмом и энерготратами
организма существует линейная
зависимость. Э\Т=20,2*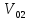 поглощенного. 20,2-среднее значение
поглощенного. 20,2-среднее значение )
является энтропия S.
S=s/T(t)
Размерность Дж*
)
является энтропия S.
S=s/T(t)
Размерность Дж*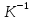
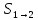 =
=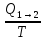
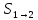 =
=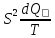
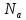 =1,38*
=1,38*
 Дж*
Дж*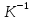
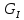 в результате совер-ния работы и
процессов диссипации энергии.Для
живых орг. справедливы выражения:
в результате совер-ния работы и
процессов диссипации энергии.Для
живых орг. справедливы выражения: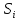 +ΔSe(общее
изм-ние энтропии=сумме изм. Энтропии
за счет процессов протекающих внутри
системы и протекающих за счет процессов
обмена с окр средой .
+ΔSe(общее
изм-ние энтропии=сумме изм. Энтропии
за счет процессов протекающих внутри
системы и протекающих за счет процессов
обмена с окр средой .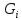 извне,но
может и отдавать другим телам,
извне,но
может и отдавать другим телам,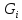 <0
поэтому ∆
<0
поэтому ∆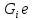 может
иметь любой знак. всегда.
может
иметь любой знак. всегда.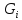 <0
∆
<0
∆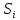 >0
>0 (
(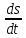 )+(
)+(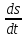 )e
–выражается производной энтропии по
времени.
)e
–выражается производной энтропии по
времени.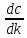 ;
grad
T=
;
grad
T=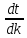 ;
gradφ=
;
gradφ=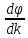 градиент векторная велечина. В случаях
биомембран используют среднее значение
gradC=
градиент векторная велечина. В случаях
биомембран используют среднее значение
gradC=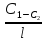 l-толщина
мембраны
l-толщина
мембраны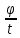
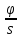
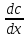 =-DSgradC
=-DSgradC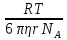
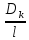 (
(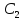 -
-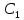 )
)
|
Билет11
Электродиффузия-дуффузия
электроически заряженных частиц
(ионов) под влиянием концептрационного
и электрического градиентов.Липидный
бислой непроницаем для ионов , они
могут проникнуть только посредством
специальных структур –ионных
каналов,которые образованы интегральными
белками.Движущей силой диффузии яв-ся
не только разность конц. Ионов внутри
и вне клетки ,но так же разность
ЭХ(электро-химического )
потенциалов,создаваемых этими ионами
по обе стороны мембраны=>диффузный
поток ионов определ-ся градиентом ЭХ
потенциала. ЭХ потенциал опрелеляет
свободную энергию иона и учитывает
все силы , способные побудить ион в
движению.µ= Зав-ть плотность потока ионов от ЭХ потенц. Определяется ур-ем Теорелла U-подвижность ионов,dµ/dx-ЭХ градиент.Подставляя выражение для ЭХ потенциала в ур-е Теорелла,можно получить урНерист-Планка с учетом 2х grad С, которые обуславливают диффузию ионов.
ϳ=D
Облегченная
диффузия гидроф
молекул.Крупные гидрофильные молекулы
(сахара, аминокислоты )перемещаются
через мембр. с помощью переносчиков.Этот
тип транспорта яв-ся диффузией
,поскольку транспорт в-ва перемещается
по gradС
без доп энергии.Другой особенностью
облегченной диффузии яв-ся Феномен
насыщения. Поток
вещ-ва,транспортируемого путем для
диффузии , растет в зависимости от
gradC
в-ва только до определенной величины.Затем
возрастание потока прекращается ,
поскольку транспортная система
полностью занята.Кинетику обл.диффузии
отображает управление Михаэлиса
Ментена. ϳ= |
Билет№12 Активный транспорт веществ. Компоненты и свойства системы активного транспорта.Na\K насос. Активный транспорт : 1)перенос веществ против физико-химического gradC; 2)с затратой энергии АТФ непосредственно на процесс переноса веществ через мембрану. Существует несколько систем транспорта ионов через плазматическую мембрану – ионный насос: 1)Na\K насос 2)Са 3)Водородный. )Na\K насос существует в плазматических мембранах всех животных и растений. Он выкачивает ионы Na из клеток, и закачивает в клетку ионы K, в результате концетрации K в клетках существенно превышает концетрацию ионов Na. Na\K насос состоит из интегральных белков мембраны, обладающий энзимными свойствами(способность гидролизовать АТФ). АТФ является основным источником запасов энергии в клетке, поэтому интегральный белок назвали Na\K АФТазы. Молекула насоса существует в 2 конформациях, взаимное преобразование стимулируется гидролизом АТФ. При повышении концетрации Na в клетке, 3 иона Na присоеденяются к белку, молекула насоса приобретает АТФазную активность. При расщеплении Na\K АТФазой молекулы АТФ неорганический фосфат присоединяется к белку. Конформация Na\K АТФазы изменяется, 3 иона Na удаляется из клетки. Затем молекула неорганического фосфата отсоединяется от насоса белка, и насос превращается в переносчик K, в результате 2 иона K попадают в клетку. Таким образом при расщеплении каждой молекулы АТФ выкачиваются 3 иона Na из клетки, и 2 иона K закачиваются в клетку. Один Na\K насос может перенести через мембрану 150-600 ионов в секунду, следствием его работы является поддержание трансмембранных градиентов Na и K
|
|
Билет№13 Концетрационный элемент Нернста. Необходимые и достаточные условия биоэлектрогенеза в уравнении Нернста.
Теория
электролетической диссоциации(ТЭД)
и концетрационный элемент Нернста
позволили понять физико-химические
основы биоэлектрогенеза. Живые ткани
содержат электролиты которые распадаются
на ионы, поэтому самой простой моделью
является источника электро-магнитной
энергии служит концетрационный элемент
Нернста. В нем растворы, соли разной
концетрации отделены мембраной имеющая
не одинаковую проницаемость для
ионов\катиона и аниона. Вследстии
существовании разности концетрации
электролит стремится диффундировать
через мембрану, но в силу неодинаковой
проницаемости мембран, одни из ионов
преодолевают ее, а другие задерживаются
ею. Происходит разобщение в пространстве
«+» и «-« зарядов, без чего невозможно
создание ЭДС. При одинаковой проникающей
способности катионов и анионов,
электрическая энергия не генерируется.
ЭДС возникающая на концетрационном
элементе образовано раствором одной
соли определяется из уравнения Нернста.
ЭДС= Таким образом, для генерации и распространения биопотенциала(биоэлектрогенеза) соблюдается 2 обязательных условия(необходимое и достаточное) 1)существование электрических концетрационных градиентов на клеточной мембране 2)наличие не одинаковой проницаемости этой мембраны для катионов и анионов на который диссоциирует электролиты в живых тканях. Источником электромагнитной энергии в любой клетке служит концетрационный элемент образованный растворами солей между цитоплазмой и межклеточной жидкостью разделенной мембраной, обладающей не одинаковой проницаемостью для катионов и анионов.
|
Билет 14
При рассмотрении
живых тканей в качестве конц элемента
необходимо учитывать диффузию через
мембрану не одного иона ,а всех.Это
обст-во учитывает ур-ие Гльдмана.
ЭДС= Двойной слой зарядов обр-ся в следствии существования разной конц.соли ,где электролит стремится диффузировать через мембрану ,но в силу неодинаковой проницаемости для катионов и анионов, некоторые ионы проходят через мембрану некоторые зац-ся . Потенциал покоя-это стационарное разность потенциалов между внешней и внутренней средой клетки ,существующая на наружней мембране клетки в невозбужденном состоянии.Внутренняя пов-ть клеточной мембраны имеет отрицательный потенциал по отношению к внеш.Реализация обязательных условий блоэлектроген на наружной мембране клетки в сост.покоя . 1.В цитоплазме преобладают К соли высших кислот, в межклеточной среде Na соли неор. Кислот Проницаемость нар. Мембраны клетки для ионной калия значительно больше чем для натрия и Cl.Такая проницаемость обусловлена наличием на наружной мембране так называемых потенциал зависимых каналов,избирательно пропускающих К,Na,Cl. Если концентрация какого либо иона внутри клетки отлична от конц.этого иона снаружи и мембрана проницаема для этого иона,возникает поток заряженных частиц через мембрану.Происходит пространственное разобщение противоположных по знаку ионов.Вследствие этого нарушается обр-ся разность потенциалов.(градиент потенциала) между наружной и внутренней средой,которая препятствует дальнейшему переносу ионов через мембрану.При установлении равновесия между конц. И элект. Градиентом выравниваются значения электро-хим. потенциалов для этого иона,которые рассчитывается по формуле Нериста
µ= Потенциальное действие называют кратковременное изменение трансмембранной разности потенциалов на наружной мембр. нервных и мышечных клеток при их возбужд. В опытах по изучению ПД исп-ют два микроэлектрода ,введенных в мышечную или нервную клетку.На 1 микроэлектрод подают импульсы ,изменяющие мембранный потенциал .Мембранный потенциал измеряют при помощи второго микроэлектрода высокоомным регистратором напряжения.
Под влиянием
раздражителей изменяются условия ЭДС
в кл.конц.элементах,что выр-ся
возникновением ПД.Проницаемость для
натрия становится гораздо выше
проницаемости аноинов.=> в момент
возбуждения
Возбужденный участок цитоплазмы изменяет свой потенциал на положительный (деполяризация),возращение к прежней поляризации-реполяризация. |
|
Билет№15 Понятие возбудимости и возбуждения. Вольт-амперные характеристики. Козбудимые и не возбудимые мембраны. Критический уровень мембранного потенциала порогового раздражителя.
Возбудимые
мембраны способны создавать не только
потенциал покоя(ПП), но и потенциал
действия(ПД), обусловлено наличием
потенциал зависимых ионных каналов.
В невозбудимых мембранах находятся
только потенциал зависимые ионные
каналы. Возбудимость- способность
возбудимой ткани изменять свои свойства
или состояния под действием раздражителей,
ее количественной мерой служит:
интенсивность порогового раздражителя.
Вольт-амперная характеристика невозбудимой мембраны линейная. При изменении мембранного потенциала (МП) от уровня ПП до 0. Электропроводность мембраны остается неизменной. С понижением абсолютного значения МП плотность ионного тока сквозб невозбудимую мембрану пропорционально уменьшается в соответствии с законом Ома. Вольт-амперная характеристика возбудимой мембраны нелинейная. При понижении трансмембранной разности потенциалов до определенного значения( не до нуля) плотность ионного тока снижается до нуля. Затем дальнейшее уменьшение абсолютного значения трансмембранного потенциала приводит к резкому нарастанию тока. Возбудимые мембраны генерируют ПД только при действии на них электрических импульсов определенной величины – пороговый и надпороговый раздражители. Минимальный импульс напряжения способный вызвать ПД на возбудимой мембране – пороговый раздражитель.
Этот импульс
сдвигает МП до определенного уровня
– критический мембранный потенциал
Величина порогового раздражителя является мерой возбудимости мембраны, тоесть ее способность генерировать ПД. Возбудимые мембраны способны генерировать ПД, процесс возникновения ПД, возбуждение. КПМ- под действием деполяризации мембраны увеличивается вероятность перехода натриевого канала в проводящее состояние. Постепенно число открытых каналов растет и входящий натриевый ток увеличивается. Одновременно увеличивается выходящий калиевый ток. Но натриевый ток растет быстрее. Уровень трансмембранной разности потенциалов при котором входящий натриевый ток начинает превышать выходящий калиевый ,носит название КМП, при к |
Билет16 Раздражимость живых тканей обусловлена тем, что в ответ на раздражители изменяется ионная проницаемость мембран,т.е. реагирование обусловлено изменением электроосмотических св-в. живой клетки. Под действием открываются Деполяризация Гиперполяризация. Раздражительный канал Выходящий анионный ток невозможен.тк.цитохоль по ср с интерстицием богаче анионами ВМС. Для них нет каналов в плазмолемме. Такие анионы покидают клетку только при повреждении. Деполяризация возникающая при этом называется потенциалом повреждения. В невозбудимых мембранах деполяризация и гиперполяризация имеют градуальный хар-р,т.е. линейная зависимость величины сдвига МП от силы раздражителя. Закон «ВСЕ ИЛИ НИЧЕГО»-если деполяризующий импульс больше порогового ,то развивается ПД, амплитуда которого не зависит от амплитуды возбуждающего импульса; если деполяр-щий импульс меньше порогового , то ПД нет. С момента установления КМП процесс развития ПД становится необратимым ,»лавинообр» открытие натриевых каналов:Входящий Na ток деполяризует мембрану->деполяризация мембраны повышает вероятность открытия Na каналов-> открываются каналы-> увел-ся входящий натриевый ток.В этот период возбудимая мембрана не чувствительна к воздействию внешних стимулов,наступает фаза абсолютной рефрактерности, длится времени ПД. Фазы рефрактерности: 1.Фаза следовой деполяризации (экзальтации) – в этот период вероятность открытия Na каналов уже высока , а МП сдвинут по направлению к КПМ. 2. Фаза следовой гиперполяризации МП сдвинут ниже уровня ПП. |
|
Билет17 Распространение возбуждения по нерву состоит из двух последоват. процессов: 1.Распространение ЭМП с затуханием 2.Ретрансляция ПД.Пусть в некоторой точке нервного волокна развился и достиг пика ПД,т.е. произошла деполяризация мембран.В месте возн. ПД потенциал внутренней стороны отрицателен. В цитозоле и интерстиции возникают ионные токи (локальные токи):между участками вор-ти мембраны с большим потенциалом (полож. Заряж) и участками пов-ти мембран с меньшим потенциалом (отрицат.заряженым) За счет этих токов потенц. участков мембран повышается. Трансмембранная разность потенциалов уменьшается по абсолютной величине ,невзбж. Участки мембраны деполяризуются.По мере удаления от точки возникновения ПД изменения трансмембранной разности потенциалов убывает по эксп.з-ну.(распр с затуханием) В тех местах ,перехват Равье,много натриевых каналов ,где сдвиг трансмембр разности выше КМП, открываются натриевые каналы и происходит развитие ПД(ретрансляция ПД) Итак , возникающий на возбудимой мембране ПД явл-ся надпороговым стимулом для определенного участка мембраны.По скольку ретрансляция ПД происходит только в перехвате Ранвье, то возбуждение как бы перепрыгивает через миелизиров. Участки мембраны: такой тип проведения возбуждения получил название сальтаторное проведение. Миелинезация обесп.повышение скорости проведения при сущ.экономии энергетических ресурсов. |
Билет 18 Синоптическая передача. Функциональный межклеточный контакт обеспечивающий переход возбуждения с одной клетки на другую, получил название синапс: а)электрический; б)химический; Электрическая синоптическая передача возможна только при очень тесном соприкосновении взаимодействующих клеток – при расстоянии между ними не более 10-20 нм. В этом случае развитие ПД на мембране одной клетки приводит за счет возникновения локальных токов и деполяризации мембраны другой клетки, которая может оказаться выше порога генерирования ПД. Большое значение для осуществления электрической передачи играет щелевые контакты (нексусы). При этом на каждой из 2 соседних мембран, регулярно расположенных коннексонов(канальные белки, обладающие высокой проходимостью ионов). Щелевые контакты регулируемы, они могут закрываться, при снижении pH или повышение концентрации иона кальция, за счет такого механизма пораженные места изолируются от остальной части синцития, распространение патологии ограничивается(инфаркт миокарда). Химическая синоптическая передача осуществляется с помощью химических веществ посредников, медиаторов, в этом случае расстояние между взаимодействующими клетками в области контакта (ширина синаптической щели больше). Электрическое поле затухает, в пределах синаптической щели, и не может деполяризовать синаптическую мембрану для медиатора. Отсюда возникает необходимость химического посредника. Деполяризация пресинапса приводит к изменению проницаемости пресинаптической мембраны для медиатора, который выбрасывается в синаптическую щель, взаимодействуетс белками-рецепторами пост.синт.мембрану,где происходит конформации белка и изм.МП. Врзбуждающий синапс-деполяриз.постостмембраны. Тормозный синапс-гиперполяриз постсинаптической мембраны.В случае возникновения возбуждающ.постсинаптического потенциала он с затуханием распр-ся по пост-синапт мембране и может вызвать возн.ПД на возбудимых уч-ках мембраны принимающий сигнал клетки, если он превышает пороговый уровень.
|
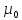 +RT*luC+ƶFȹ
+RT*luC+ƶFȹ
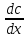 -uƶFC
-uƶFC Ионные
каналы мембраны представляют собой
интегральные белки мембраны ,которые
обр-ют отверстие в мембране,заполненных
водой. В плазмолемме обнаружен ряд
ионных каналов, которые х-ются высокой
спецефичностью ,допускающих перемещение
протока одного вида ионов.Существуют
Na,Cl
каналы,каждый из них имеет селективный
фильтры,который способен пропускать
только определенные ионы.Проницаемость
ионных каналов
может измениться благодаря наличию-ворот
определенных групп атомов в составе
белков ,форм канал. Конформационные
изм-ния ворот при изменении ЭХпотенциала
или действием спецефических химических
в-ввыполняющих сигнальную функцию.
Ионные
каналы мембраны представляют собой
интегральные белки мембраны ,которые
обр-ют отверстие в мембране,заполненных
водой. В плазмолемме обнаружен ряд
ионных каналов, которые х-ются высокой
спецефичностью ,допускающих перемещение
протока одного вида ионов.Существуют
Na,Cl
каналы,каждый из них имеет селективный
фильтры,который способен пропускать
только определенные ионы.Проницаемость
ионных каналов
может измениться благодаря наличию-ворот
определенных групп атомов в составе
белков ,форм канал. Конформационные
изм-ния ворот при изменении ЭХпотенциала
или действием спецефических химических
в-ввыполняющих сигнальную функцию.
 КМ-константа Михаэлиса равна конц-ции
в-ва вне плотности потока равна половине
максимальной.
КМ-константа Михаэлиса равна конц-ции
в-ва вне плотности потока равна половине
максимальной.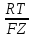 ln
ln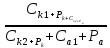
 ln
(фор.дописать)
ln
(фор.дописать)
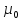 +RTlnC+ƶFȹ
+RTlnC+ƶFȹ >
>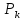 >
>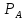 Потенциал инверсии равен превышению
на абсолютной величине ПДнад ПП.
Потенциал инверсии равен превышению
на абсолютной величине ПДнад ПП.
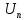 =
=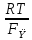 ln
ln

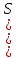 )
ԑ(возбудимость)=
)
ԑ(возбудимость)=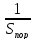
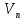 =│ПП│-│КМП│
=│ПП│-│КМП│