

Биомы, Гл
.9.pdfвысотой до 10 м образуют бамбуки (Chus-quea quila, Ch. colihue), древовидные папоротники (Blechnum magellanicum), веч- нозеленые кустарники мирта, барбариса и др. В наземном покрове – красная водяника, ясколка, папоротники и обильные мхи. В этих летнезел¸ных лесах много торфяных болот.
Лесная растительность сильно нарушена. Особенно интенсивно сводится лес на восточных, защищенных от ветра склонах хребтов и на равнинах. Земли после вырубки и выжигания подвергаются сильной эрозии, на склонах образуются оползни. Рас- чищенные от леса участки используются главным образом под пастбища и посевы кормовых трав для крупного рогатого скота.
Животный мир Патагонии сохранился преимущественно в горах. Он неотделим от Неотропического царства. Характерные представители фауны Патагонско-Чилийс- кой области показаны на рис. 9.113.
Юг Австралии, остров Тасмания и Новая Зеландия южнее 38î ю. ш. располагаются в пределах умеренно-теплой зоны. Климат влажный, океанический. Средняя температура самого холодного месяца 5 – 8 îС; лето прохладное – 14 – 18 îС. Годовое количе- ство осадков превышает 1000 мм; распределяются они по сезонам равномерно. Благодаря теплой зиме вегетационный период продолжается практически круглый год.
В растительном покрове много вечнозеленых видов, что придает биомам зональных типов субтропический облик. Их характерные черты наиболее ярко выражены в Новой Зеландии. Отличительной чертой растительности являются леса из южного бука (Nothofagus), аналогичные ЧилийскоПатагонским.
Это дает основание ботаникам относить Новозеландскою область к Голантарктическому царству. Зоогеографы выделяют Новозеландскую область в составе Австралийского царства (см. рис. 9.72).
9.2.5.Степи
Âумеренно-теплых зонах по мере усиления засушливости климата лесные биомы сменяются травянистыми безлесными сообществами. В травостое господствуют многолетние зимостойкие, способные переносить длительную засуху злаки и ксерофитное разнотравье. Этот тип растительности в Северном полушарии в Евразии называют степью, в Северной Америке – прерией, в Южном полушарии в Аргентине – пампой, в Новой Зеландии – туссоком. Учи- тывая господствующую роль злаков в биомах этого типа, употребляют и общее название – злаковники. Обращает внимание, что такие роды злаков, как ковыль (Stipa), овсяница (Festuca), мятлик (Poa), житняк (Agropyrum) и др., доминирующие в растительном покрове злаковников, имеют биполярное распространение.
Северное полушарие. От венгерских пушт через всю центральную часть Евразии до Алтая и далее через Забайкалье в Монголию и Северо-Восточный Китай простираетсяПалеарктическая степная область Голарктического царства. Граница степных биомов с более северными лесными не резкая. Переходом служит лесостепь, представляющая собой мозаичный макрокомплекс, в котором массивы леса и пространства луговых степей резко отграничены друг от друга. Этим лесостепь отличается от саванны, где отдельно стоящие деревья естественно вкраплены в травянистый покров.
Биомы степей геологически сравнительно молоды. В пределах современного ареала они стали формироваться в основном уже в послеледниковое время. В тече- ние ледникового периода в так называемой перигляциальной зоне по окраине ледникового щита располагалась обширная тундростепь.
Степная флора, однако, существовала и ранее. В доледниковое время степные формации уже отмечались в пределах древнего Средиземья (в Передней и Средней Азии и в Иране); а также в горах Карпат, Балканского полуострова, на Кавказе и в Средней Азии. Все эти центры дали основную мас-
226
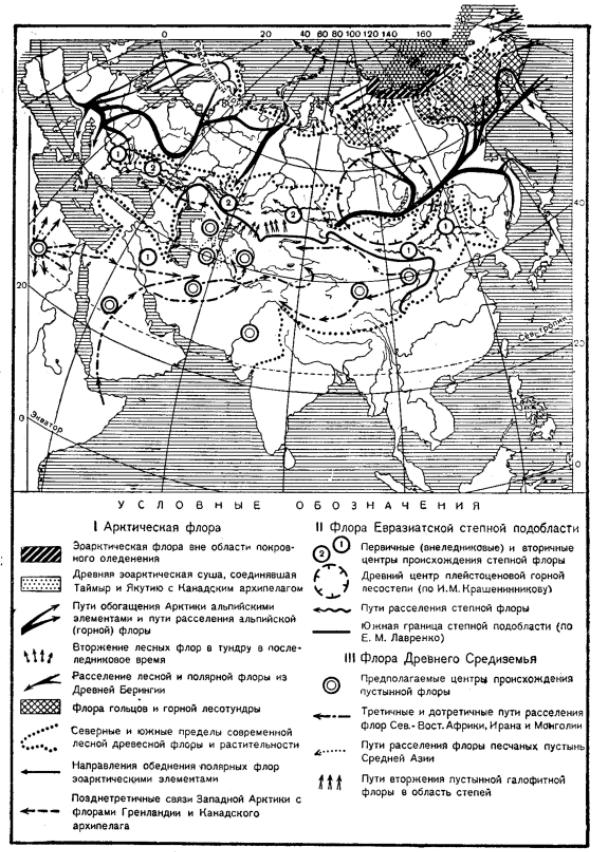
Рис. 9.114. Схема путей миграций и центров распространения флор степей, пустынь и тундр Евразии, по В. В. Алехину и В. С. Говорухину
227

су ксерофитной флоры современных степей Западной Евразии (рис. 9.114).
Степную растительность образуют |
|
|
|
виды, сформировавшиеся в основном в теп- |
|
|
|
лых и сухих условиях средиземноморского |
|
|
|
климата. С наступлением похолоданий в |
|
|
|
плейстоцене эта ксеротермофитная расти- |
|
|
|
тельность трансформировалась в криоксе- |
|
|
|
рофитную перигляциальную. Северные бо- |
|
|
|
лее влаголюбивые виды (широкотравье, кор- |
Рис.9.116. Широтно-зональная смена растительного |
||
|
|||
невищные луговые мезофитные злаки и осо- |
покрова степей, по В. Г. Мордковичу: |
||
ки северных степей) проникли в степи из |
А – северная луговая степь; Б – типичная ковыльная |
||
лесной зоны. |
степь;В – южная типчаково-ковыльная степь |
||
Огромная протяженность евразиатских |
ния масс плотного и тяжелого воздуха. На- |
||
степных ландшафтов с севера на юг и с за- |
помним, что в умеренном поясе над Атлан- |
||
пада на восток определяет зональные и про- |
тическим океаном располагается баричес- |
||
винциальные изменения растительных со- |
кий центр низкого давления. Воздушные |
||
обществ. С запада на восток выделяют про- |
массы устремляются из Азии в западном на- |
||
винции: Европейско-Казахстанскую, Забай- |
правлении. Отрог изобар высокого давле- |
||
кальскую, Центральноазиатскую. С севера |
ния, тянущийся на запад от Сибирского ан- |
||
на юг степи делятся на подзоны. Так, в Ев- |
тициклона, был впервые описан в начале |
||
ропейско-Казахстанской провинции выде- |
ХХ века климатологом А. И. Воейковым и |
||
ляют подзоны северных луговых, типичных |
получил название “оси Воейкова”. Эта ось |
||
и южных сухих степей (рис. 9.115). По мере |
служит ветроразделом: к северу от нее дуют |
||
увеличения засушливости климата меняет- |
теплые, влажные, несущие осадки западные |
||
ся облик растительности (рис. 9.116). |
ветры (западный перенос); к югу преобла- |
||
|
дают сухие и холодные северо-восточные |
||
|
и восточные ветры. По мере продвижения |
||
|
запад воздействие азиатского антициклона |
||
|
ослабевает. Степная зона Восточной Евро- |
||
|
пы доступна теплым и влажным воздушным |
||
|
массам, приходящим с Атлантики. Средние |
||
|
температуры января в Молдавии -4 îÑ (àá- |
||
|
солютный минимум до -25 îÑ); â Òóâå, íà- |
||
|
ходящейся во власти зимнего антициклона, |
||
|
äî |
-35 îС (абсолютный минимум до |
|
|
-50 îÑ). |
||
|
|
Весной Сибирский антициклон осла- |
|
Рис. 9.115. Деление степной зоны на подзоны, |
бевает, а затем исчезает. На его месте вслед- |
||
по А. А. Чибилеву: |
ствие прогревания земной поверхности и |
||
1 – лесостепь; 2 – северная луговая степь; |
нижних слоев атмосферы устанавливается |
||
3 – типичная степь; 4 – южная степь |
|||
область низкого давления. Значения темпе- |
|||
|
|||
|
ратуры июля мало различаются по мере дви- |
||
Усиление континентальности климата |
жения по долготе: в Придунайской низмен- |
||
в степной зоне связано с действием Сибир- |
ности 23 îС, на Южном Урале 22 îÑ, â Çà- |
||
ского антициклона. Мощный барический |
байкалье 20 îÑ. |
||
центр устойчивого высокого давления фор- |
|
Важным фактором формирования степ- |
|
мируется осенью и зимой над просторами |
ных биомов является количество и ритми- |
||
Центральной Азии вследствие активного |
ка выпадения атмосферных осадков. В ев- |
||
выхолаживания территории и формирова- |
ропейской лесостепи выпадает до 500 мм, |
||
228

к югу и востоку количество осадков снижается – в Туве 215 мм. Испаряемость в Восточноевропейском секторе около 1000 мм.
Сезонный ход выпадения |
|
|
осадков определяется в Европе за- |
|
|
падным переносом, в Центральной |
|
|
Азии муссонами. В европейских |
|
|
степях осадки выпадают в основ- |
|
|
ном весной и осенью; летом царит |
|
|
засуха, процессы жизнедеятельно- |
|
|
сти замирают. В степных областях |
|
|
Центральной Азии наблюдается |
|
|
летний пик увлажнения на фоне за- |
|
|
сушливых и холодных весны и осе- |
|
|
ни (рис. 9.117). Здесь под действи- |
|
|
ем летнего муссона дожди выпада- |
|
|
ют в течение 1 – 1,5 месяцев (в |
|
|
июле – августе). Поскольку в этот |
|
|
же период наблюдается и максимум |
|
|
тепла, интенсивность жизнедея- |
Рис. 9.118. Карта типов почв степной области Восточной Европы и |
|
тельности высокая. |
прилегающих лесных районов, по Г. Вальтеру: |
|
Выдающимся феноменом |
1 – подзолистые почвы; 2 – луга и болота; 3 – карбонатное засоле- |
|
степных биомов являются черно- |
ние; 4 – серые лесные почвы; черноземы: 5 – северные; |
6 |
земы – самые богатые почвы на |
– мощные; 7 – обыкновенные; 8 – южные;9 – каштановые и бурые |
|
пустынно-степные почвы; 10 – песчаные дюны; 11 – каменистые |
||
планете. Классическая область их |
почвы. п – поды (бессточные степные блюдца); с – засоленные |
|
распространения – причерномор- |
почвы (солончаки) |
|
ские и приазовские степи. Карта |
|
|
типов почв степной области Восточной Ев- |
ной структуре степных биомов в целом (рис. |
ропы и прилегающих лесных районов дает |
9.118). |
наглядное представление о пространствен- |
Высокая продуктивность травянистой |
|
растительности, засушливое лето и известь- |
|
содержащие почвообразующие и материн- |
|
ские породы (л¸сс, мел, известняк) – вот три |
|
определяющих условия формирования ти- |
|
пичного чернозема. |
|
В структуре растительных сообществ |
|
мощно развиты как надземная, так и под- |
|
земная части (рис. 9.119). Степные травы |
|
ежегодно продуцируют десятки тонн зеле- |
|
ной и корневой массы на гектар. Эта масса |
|
потребляется растительноядными живот- |
|
ными; значительная ее часть отмирает и раз- |
|
лагается редуцентами. Накопление гумуса в |
|
черноземах является следствием неполно- |
|
го цикла деятельности микроорганизмов, |
|
которые из-за летней засухи и зимних мо- |
|
розов успевают осуществить лишь первые |
Рис. 9.117. Сезонная ритмика выпадения атмосферных осад- |
стадии трансформации органического веще- |
ков в степных биомах Восточной Европы и Центральной |
ства. Это приводит к резкому доминирова- |
Азии, по В. Г. Мордковичу |
|
229
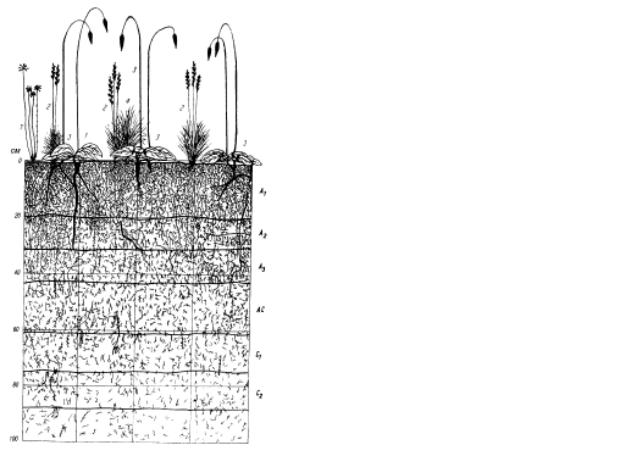
Рис. 9.119. Вертикальная структура степного сообщества, по М. С. Шалыту:
1 - Euphorbia segueriana; 2 – Festuca sulcata;
3 – Salvia nutans, 4 - Stipa lessingiana
нию в черноземах процессов гумификации органических остатков над процессами их минерализации. Известьсодержащие материнские породы способствуют связыванию гуминовых кислот в труднорастворимые гуматы кальция. Таким образом, осуществляется главный процесс формирования черноземов – гумусонакопление.
По мере продвижения на юг условия для образования черноземов в связи с повышением температуры и усилением засушливости климата становятся все менее благоприятными: уменьшается мощность гуму- сово-аккумулятивного горизонта и иллювиального горизонта (горизонта вмывания). Последний формируется под действием нисходящего потока веществ. Поднимается граница непромачиваемого почвенного горизонта, в котором наличие карбонатов устанавливается с помощью соляной кислоты, – от капли кислоты начинается бурное
выделение углекислого газа, почва “вскипает”.
Рис. 9.120. Широтно-зональная смена почв, по В. Г. Мордковичу:
1 – гумусово-аккумулятивные горизонты; 2 –ил- лювиальные горизонты; 3 – материнская порода; 4 – потолок вскипания карбонатов под действием соляной кислоты
Подзональные различия степных биомов находят отражение в почвенном покрове: от луговых степей к опустыненным последовательно сменяют друг друга мощные, обыкновенные и южные черноземы, темно- и светло-каштановые почвы (рис. 9.120).
Богатство почв предопределило судьбу степных биомов: почти повсеместно они превращены в пахотные угодья. Некоторое представление о первозданной степи дают картины художников, произведения поэтов и писателей XIX века (рис. 9.121) и заповедники.
Выше уже были названы подзоны восточноевропейских степей. При движении с севера на юг отмечаются следующие изменения в растительном покрове: 1) густой травостой луговых степей сменяется разреженным на юге; 2) красочность степей уменьшается; 3) на севере преобладают многолетники, к югу значительно усиливается роль однолетников; 4) широколистные злаки к югу уступают место злакам узколистным – ковылям, типчаку и др.; 5) видовая насыщенность (число видов на определенную площадь) убывает от 50 – 80 видов на
230

(Stipa lessingiana), ковыль-тырса (S. capillata); кроме того типчак (Festuca valesiaca), келерия (Koeleria cristata); полыни (Artemisia).
“Курской растительной аномалией” назвал в начале ХХ века профессор В. В. Алехин степи под Курском. Но эта “аномалийность” связана не с тем, что нетипична растительность, а с тем, что здесь сохранились в условиях сплошной распашки одни из немногих нетронутых целинных участков лу-
говой степи. На этих землях организован Центрально-Черноземный государственный заповедник им. В. В. Алехина.
231

Рис. 9.123. Характерные представители фауны Европейско-Казахских степей. Рис. В. А. Ватагина, по Н. А. Бобринскому:
1 – обыкновенный хомяк; 2 – крапчатый суслик; 3 – степная мышевка; 4 – слепыш; 5 – степная пеструшка; 6 – заяц-русак; 7 – дрофа; 8 – большой тушканчик; 9 – байбак; 10 – степная пищуха; 11 – степной жаворонок; 12 – черный жаворонок; 13 – краснощекий суслик; 14 – стрепет; 15 – сайгак; 16 – полоз желтобрюхий
232

Рис. 9.124. Характерные представители фауны Забайкальских степей. Рис. В. А. Ватагина, по Н. А. Бобринскому:
1 – копытка; 2 – даурская пищуха; 3 – корсак; 4 – манул; 5 – тарбаган; 6 – заяц-толай; 7 – дзэрэн; 8 – даурский суслик; 9 – монгольская песчанка; 10 – даурский цокор; 11 – монольский жаворонок; 12 – монгольская ящурка; 13 – тушканчик-прыгун; 14 – черный журавль
233
Степная растительность заповедника характеризуется богатством видового состава – до 220 видов трав – и большим числом сезонных аспектов: с начала весны до осени их насчитывают до одиннадцати. Общая картина чередования весеннелетних аспектов в луговой степи изображена на рис. 9.122.
Луговые степи необыкновенно красочны. В течение веге-
тационного периода они постоянно изменяют свой облик. Ранней весной степь бурая, но уже в апреле, как по волшебству, раскрывает нежные лиловые бутоны сон-тра- ва, за ней спешит адонис весенний. Его крупные солнечно-желтые цветки появляются еще до развития листьев. Во время цветения адониса степь постепенно одевается в зеленый наряд. Зацветают незабудки, ирис безлистный, ветреница лесная. Начало лета знаменует цветение лугового шалфея. Наиболее красочной бывает степь в это время. Но вот зацветает перистый ковыль. Все остальные цветущие виды разнотравья скрываются под его пушистыми перьями. В это время степь особенно прекрасна, похожа на седое бескрайнее море. Проходят дни, постепенно степь одевается ярко-желтыми соцветиями подмаренника. Живокость клиновидная и чемерица черная – последние цветущие растения уходящего лета.
Животный мир евразиатских степей так же, как растительность, обладает своими подзональными и провинциальными особенностями. Например, различия в фауне Европейско-Казахстанских и Забайкальских степей можно увидеть, сравнив рис. 9.123 и 9.124.
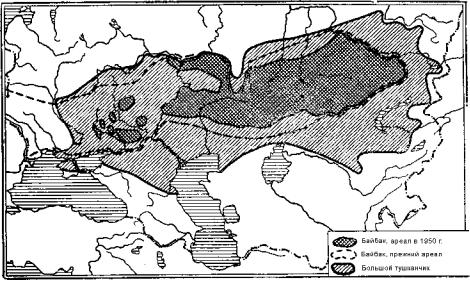
Ограничимся краткой характеристикой животного мира Европейско-Казахстанской степной провинции. Ее границы хорошо очерчивают ареалы байбака и большого туш-
канчика (рис. 9.125). Фауна провинции представляет собой пестрый комплекс, переходный от биомов широколиственного леса к азиатским пустыням. Тем не менее, он имеет эндемичное ядро, в состав которого входят несколько видов сусликов, байбак, слепыш, большой тушканчик, обыкновенный хомяк, степная мышовка. К этой же группе относится антилопа сайгак. Из наиболее характерных птиц следует назвать стрепета, дрофу, малого журавля, степного луня, степного орла; из пресмыкающихся степную гадюку, разноцветную ящурку.
Для естественных ландшафтов открытых степных равнин характерны крупные стадные копытные и грызуны. Отсутствие естественных укрытий обусловило то, что степные копытные обладают быстрым бегом и хорошим зрением, а грызуны укрываются в норах. Суровая зима заставляет многих зверей и птиц откочевывать в более южные районы.
Следует подчеркнуть, что названные группы животных не только потребляют растительную массу, но и активно воздействуют на почвенно-растительный покров, разбивая дернину и роя многочисленные норы. Высокая биопродуктивность растительности составляет основу цепей питания в степных экосистемах (рис. 9.126).
Наблюдения в заповедниках показали, что умеренные нарушения являются необходимыми для устойчивого развития степ-
234
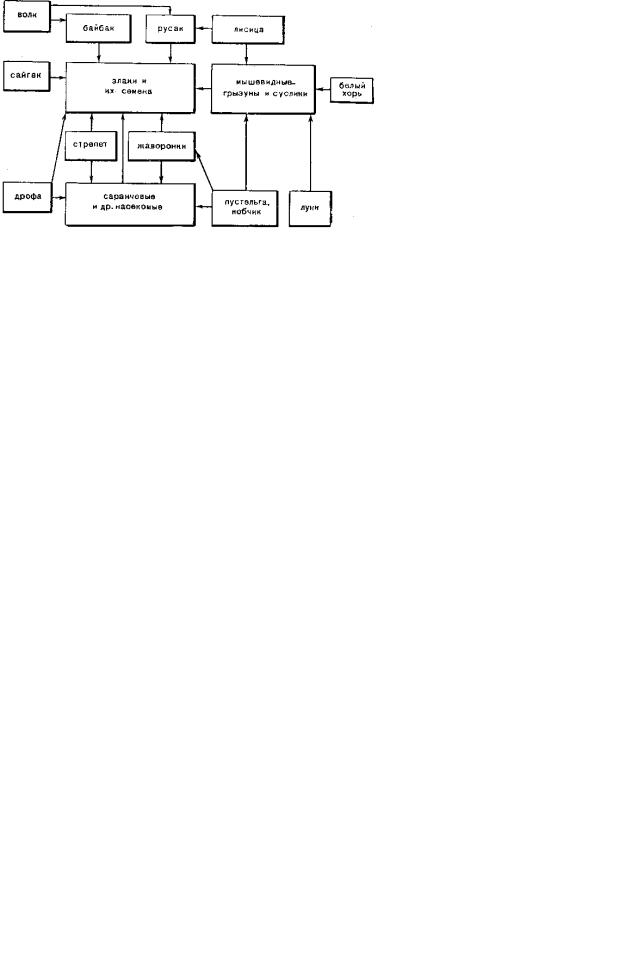
Рис. 9.126. Цепи питания в степных экосистемах, по Н. А. Бобринскому
ных сообществ: благодаря им появляются открытые участки по- чвы, доступные для семенного возобновления. Если же биопродуктивность растений не регулируется, накапливается мощный слой ветоши, дернины злаков уплотняются, так что семенное возобновление становится невозможным, многие виды не выдерживают конкуренции и биоразнообразие сообществ резко падает. Следует сказать, что в естественных условиях численность животных и биопродуктивность растительности саморегулируются и степные биомы развиваются устой-чиво.
Степные биомы Европейско-Ка- захстанских степей испытали особенно сильное воздействие человека. В Восточной Европе практически все степи распаханы, рассечены густой сетью дорог, заняты городами и промышленными предприятиями. Вместе с растительным покровом исчезли
Рис. 9.128. Ареалы прерий, по Т. Н. Гордеевой и О. С. Стрелковой:
Прерии: 1 – настоящая высокотравная восточная и южная; 2 – низкотравная и разнотравная; 3 – северные из овсяницы (Festuca); 4 – северо-западная; 5 – пустыни и полупустыни; 6 – территории с преобладанием засоленных почв, растительный покров несомкнутый
многие виды растений и животных, у других резко сократились ареалы.
Неарктическая область прерий Голарктического царства в
Северной Америке располагается вдали от атлантического океанического сектора, занятого лесными биомами (см. рис. 9.110). Данные об изменениях климата, растительности и почв прерий с востока на запад к подножью Скалистых гор отображены на профиле (рис. 9.127).
235