

Биомы, Гл
.9.pdfДоминантами растительного покрова являются дерновинные злаки, на востоке – высокие, а на сухом западе – низкие. Общая картина ареалов прерий изображена на карте (рис. 9.128).
В высокозлаковой или настоящей прерии господствуют дерновинные злаки, образующие травостой высотой 0,5 – 1 м. К числу доминантов относятся: ковыль (Stipa spartea),овсяница (Festuca scabrella), житняк (Agropyrum smithii), бородач, или грама
(Bouteloua curtipendula), тонконог (Koeleria
Рис. 9.130. Стадо степных бизонов
cristata); обильно разнотравье. Эти прерии почти сплошь распаханы.
Низкозлаковая прерия покрывает пространства Великих равнин. Здесь абсолютное господство переходит к плотнодерновинным злакам: бизоновой траве (Buchlo¸ dactyloides), траве грама (Bouteloua gracilis) (рис. 9.129). Эти прерии используются под пастбища или орошаемое земледелие.
Рис. 9.129. Доминанты низкозлаковых прерий:
а – бизонова трава (Buchlo¸ dactyloides); б – трава грама (Bouteloua gracilis)
Прерии относятся к обширной Сонорской области Неарктики, включающей широколиственные леса и субтропические биомы (см. рис. 9.68). Символом животного мира мог бы служить степной бизон, поголовье которого до колонизации Северной Америки европейцами, вероятно, достигало 50 – 60 млн. (рис. 9.130). Индейцы не могли нанести ему существенного урона, так как охотились с копьями или луком и стрелами. С 1872 по 1874 год белыми было убито более 5 млн бизонов, а к концу 1889 г. их осталось всего около тысячи. Для степного бизона роковой оказалась постройка транс-
Рис. 9.131. Охота на бизонов с поезда трансконтинентальной железной дороги в конце XIX века
континентальной железной дороги. Отстрел животных происходил прямо с проходящих поездов (рис. 9.131).
Южное полушарие.Основные площади злаковников располагаются в Южной Америки и Новой Зеландии.
Пампа – южноамериканский вариант субтропических степей занимает равнины Параны – Ла-Платы (рис. 9.132). Рельеф плоский, равнинный.
Климат субтропический. Зима теплая, температура самого холодного месяца 8 – 12 îС; летние температуры 22 – 25 îС. Годовое количество осадков возрастает от 500
– 600 мм на западе до 1000 мм у атланти- ческого побережья. Испаряется влаги несколько больше, чем выпадает. Два-три раза в течение каждых 10 лет случаются очень засушливые годы. Основная почвообразующая порода в пампе – л¸сс. На л¸се формируются черноземовидные почвы.
Безлесие пампы – явление во многом вторичное, возникшее еще до прихода европейцев, в результате выжигания растительности индейцами. К причинам безле-
236
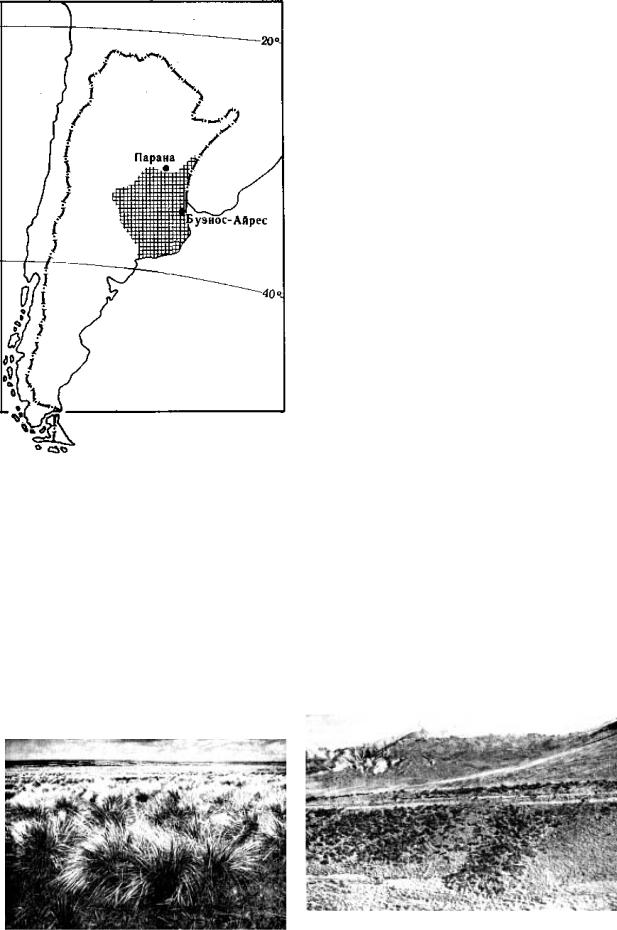
Рис. 9.132. Ареал пампы в Южной Америке, по Г. Вальтеру
сия относятся также периодические засухи, слабая дренированность и засоление грунтов.
В настоящее время - это наиболее густо населенные сельскохозяйственные районы Южной Америки. Первичная растительность пампы практически уже не существует. Однако в результате ее реконструкции установлено, что доминантами являлись злаки, многие рода которых характерны для лугов, степей и прерий Северного полушария: ковыль (Stipa), мятлик (Poa),
Рис. 9.133. Пампа с ковылем (Stipa brachychaeta), по Г. Вальтеру
аристида (Aristida), перловник (Melica), костер (Bromus) и др. (рис. 9.133); кроме того, для пампы характерны свои родаPaspalum,
Ischaemum, Cortaderia, Gynerium и др. Разнотравье представлено лупином (Lupinus), горошком (Vicia), вербеной (Verbena), портулаком (Portulaca), видами паслена.
Фауна пампы относится к Патагонско- Чилийской области Неотропического царства (см. рис. 9.113). В прошлом здесь обитали стада пампасского оленя и гуанако. В настоящее время многочисленны лишь грызуны - вискаша; встречаются броненосец, страус нанду.
В Новой Зеландии злаковники известны под названием туссок. Они распространены на Южном острове в дождевой тени Южных Альп от морского побережья до высоты 1500 м. Климат умеренно-теплый. Годовое количество осадков 600 – 500 мм. Почвы буро-серые.
Естественный растительный покров представлен злаковыми кочкарниками. Доминирующие рода – овсяница (Festuca, F. novae-zelandiae), мятлик (Poa, P. caespitosa), житняк (Agropyrum, A. scabrum) – характерны для степей Голарктического царства, т. е. имеют биполярный ареал. Это так называемый низкий туссок (рис. 9.134). Всего в группу формаций низких злаковников входит около 40 видов злаков и 35 видов сложноцветных. На влажных торфянистых почвах произрастает высокий туссок, образованый дантонией (Danthonia raoulii). Флористически эта формация беднее, так как высокие и плотно расположенные дернины дантонии подавляют другие виды.
Рис. 9.134. Туссоковый злаковник на о. Южный, Новая Зеландия, по Г. Вальтеру:
Доминирует овсяница (Festuca novae-zelandicae)
237
Со времен первых переселенцев злаковники Южного острова использовались как пастбища, поэтому девственных, нетронутых европейцами и домашними животными злаковых биомов не сохранилось.
9.2.6.Внетропические пустыни
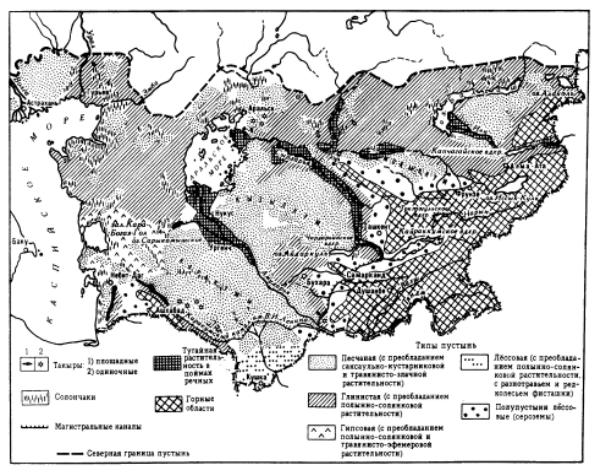
Âконтинентальном секторе Евразии к юго-востоку от зоны степей располагаются пустыни. Они занимают обширные пространства умеренного пояса Средней и Центральной Азии (рис. 9.135). На юге они смыкаются с пустынями жаркого пояса.
Переход от пустынь жаркого пояса к внетропическим пустыням происходит постепенно. Он связан прежде всего с понижением зимних температур. Устойчивый морозный период определяет своеобразие биомов. Границей, разделяющей тропические и внетропические пустыни, могут служить пределы возделывания финиковой пальмы, которая не переносит длительных заморозков.
Общие сведения о рельефе пустынь, формировании пустынных биомов, адаптациях растений и животных к жизни в условиях высоких температур и дефицита влаги рассмотрены при описании пустынь жаркого пояса (см. раздел 9.1.3). Остановимся на некоторых особенностях биомов пустынь Средней Азии и Казахстана.
Южные степи и северные пустыни объединены полосой динамического контакта, выделяемой в качестве полупустыни. По мере продвижения на юг дерновинные злаки постепенно уступают место пустынным полукустарникам: видам полыни (род
Artemisia, подроды Seriphidium и Dracunculus), прутняку (Kochia prostrata), разнообразным солянкам и т. п. Следует под- черкнуть, что наряду с полынью семейство маревые, или лебедовые (Chenopo-diaceae), куда входят все виды, объединяемые термином “солянки”, является ландшафтообразующим. Ареал саксаула (род Haloxylon, сем.
Chenopodiaceae) довольно точно очерчивает границы пустынь Средней Азии и Казахстана (рис. 9.136).
Пустынные ландшафты Туранской низменности простираются на 10 – 12î по широте. На этом протяжении зональные изменения термических условий и режима увлажнения приводят к формированию двух подзон – северных и южных пустынь. Климатическими условиями
образования внетропических пустынных биомов, общими для пустынь жаркого пояса являются: большая солнечная радиация, исключительная засушливость всего вегетационного периода, высокие летние температуры (25 – 30 îС), малое количество ат-
Рис. 9.136. Ареал рода саксаул, по “Жизнь растений”
238
мосферных осадков (в среднем 100 – |
ной стадии. Значительное участие в почво- |
150 ìì). |
образовании принимают водорастворимые |
Испаряемость в 10 – 25 раз превышает |
соли – карбонаты, сульфаты и поваренная |
сумму выпадающих осадков. Зимние темпе- |
соль. Они переходят в почву из сильно ми- |
ратуры отрицательные -10, -15 îÑ, ëèøü íà |
нерализованных материнских пород или |
юге Туранской низменности они несколько |
поднимаются по капиллярам из грунтовых |
âûøå 0 îС; абсолютные минимумы на се- |
вод. В то же время в пустынных почвах |
вере Казахстана достигают -50 îÑ, â Òàø- |
очень мало гумуса (менее 1 %). Это связано |
кенте -30 îÑ. |
с тем, что растительность бедна, и в почву |
В жизни пустынь в условиях дефицита |
поступает небольшое количество органи- |
атмосферных осадков большое значение |
ческих остатков, которые к тому же быстро |
имеют подземные воды, в частности грун- |
минерализуются. Пестрота почво-грунтов, |
товая верховодка. В пустынях, сформиро- |
разная степень засоления, условия увлаж- |
ванных на месте древних аллювиальных |
нения– причины комплексности раститель- |
равнин, они образуют сплошное зеркало. |
ного покрова. |
Грунтовые воды засолены, часто залегают |
В основе классификации пустынь |
неглубоко (3 - 4 м) и доступны для корней |
Средней Азии и Казахстана лежит характер |
растений; местами располагаются линзы |
поверхностных отложений и почвогрунтов. |
пресной верховодки. |
Основными типами пустынь являются: пес- |
Засушливый климат обусловил особый |
чаные, глинистые, гипсовые, лессовые; сво- |
тип почвообразования: для южных пустынь |
еобразные природные комплексы представ- |
характерны различного типа сероземы, для |
лены такырами, солончаками и поймами |
северных – серо-бурые почвы. На огромных |
рек; особый тип ландшафтов связан с гора- |
территориях песчаных пустынь почвообра- |
ìè (ðèñ. 9.137). |
зовательный процесс находится в началь- |
|
Рис. 9.137. Пустыни Средней Азии и Казахстана, по: “Пустыни”
239

В тесной связи с почвогрунтами нахо- |
На нем отчетливо видны гряды песков, |
|
дится растительность. Выделяются следую- |
заросшие псаммофитами, и солончаковая |
|
щие типы пустынной растительности (вне |
депрессия с сообществами галофитов. |
|
речных пойм): псаммофитный древесно- |
Фауна пустынь Средней Азии и Казах- |
|
кустарниковый на песках (Haloxylon |
стана относится к Арало-Каспийской обла- |
|
persicum, H. aphyllum, Calligonum caput- |
сти Палеарктики (рис. 9.139). Она отлича- |
|
medusae, C. setosum, Salsola richteri, |
ется своими специфическими особенностя- |
|
Ammodendron conollyi, Ephedra strobilacea è |
ми и вместе с тем имеет переходный харак- |
|
др.); гипсофильный мелкокустарничковый |
тер между сахара-средиземноморской и цен- |
|
на щебнистых и каменистых третичных |
тральноазиатской фаунами. |
|
плато (Salsola arbuscula, S. rigida, S. |
Адаптации растений и животных к |
|
gemmascens, S. laricifolia, Anabasis salsa, |
жизни в условиях пустыни вырабатывались |
|
Hammada leptoclada); эфемерово-эфемеро- |
на протяжении более 100 млн. лет, с мело- |
|
идный на л¸ссах; полукустарниковый по- |
вого и третичного периодов. Они шли по |
|
лынный на суглинках (Artemisia semiarida, |
пути приспособления к засухе, жаре, боль- |
|
A. kemrudica, Kochia prostrata, Eurotia |
шой амплитуде температур, бедным и засо- |
|
ceratoides с участием многолетних злаков, а |
ленным грунтам, к сыпучим пескам и |
ò. |
также эфемеров и эфемероидов); водорос- |
п. Главное направление адаптаций у расте- |
|
лево-лишайниковый на такырах со своеоб- |
ний привело к образованию многочислен- |
|
разными группировками пустынных ли- |
ной группы ксерофитов (см. табл. 9.2). |
|
шайников и синезеленых водорослей (Пет- |
Основными приспособительными |
|
ðîâ, 1973) . |
признаками ксерофитов являются особен- |
|
В качестве иллюстрации зависимости |
ности формирования их листа, ведущие к |
|
растительности от почвогрунтов приведен |
уменьшению размеров листовой пластин- |
|
аэрофотоснимок песчаной пустыни Караку- |
ки – микрофилии и афилии (Василевская, |
|
ìû (ðèñ. 9.138). |
1954). Продукция надземной части пустын- |
|
|
ных растений ничтожна по сравнению с |
|
|
продукцией подземной: на корневую систе- |
|
|
му приходится до 95% всей фитомассы. Кор- |
|
|
ни очень глубоко проникают в землю, дос- |
|
|
тигая капиллярной каймы грунтовых вод на |
|
глубине до 20 – 25 м.
Весьма своеобразны приспособления псаммофитов – растений, живущих на рыхлых, иногда даже подвижных песках. Их корни протягиваются по склонам барханных цепей на 25 – 30 м и уходят в глубину на 2- 2,5 м, достигая капиллярного подпора грунтовых вод и обеспечивая растения достаточным количеством влаги. Во время передвижения песков ветер выдувает из-под корней грунт, они частично обнажаются, но не гибнут, так как хорошо защищены от палящих лучей солнца. Засыпанные песком кусты не погибают. Они дают массу придаточ- ных корней, способствующих закреплению песка (рис. 9.140).
Плоды и семена многих псаммофитов
Рис. 9.138. Грядовый рельеф песков с сообществами псам- имеют приспособления для переноса вет-
мофитов; серое овальное пятно (а) – солончаковая деп- ром (рис. 9.141). У песчаной акации они рессия с сообществами галофитов. Аэрофотоснимок
240

Рис. 9.139. Характерные представтели фауны Арало-Каспийских пустынь. Рис. В. А. Ватагина, по Н. А. Бобринскому:
1 – тонкопалый суслик; 2 – саксаульная сойка; 3 – гребнепалый тушканчик; 4 – ушастая круглоголовка; 5 – цинковый геккон; 6 – варан; 7 – каракал; 8 – белобрюхий рябок; 9 – барханный кот; 10 – сырдарьинский фазан; 11 – степная черепаха; 12 – полуденная песчанка; 13 – ушастый еж; 14 – джейран; 15 – пегая землеройка; 16 – кулан
241

увеличивает сосущую силу корней и уменьшает транспирацию.
Одной из характернейших черт приспособления растений к суровым условиям пустынь является их способность уменьшать продолжительность вегетационного периода или впадать в состояние анабиоза. Это связано с периодичностью в годовом ходе выпадения атмосферных осадков, их малым количеством, летней жарой и зимни-
ми холодами. В резуль-
Рис. 9.140. Ярусная корневая система Ephedra strobilacea, сформировавшаяся в тате этого в пустынях
результате засыпания растения песком, по Н. Т. Нечаевой
сформировались две
имеют пропеллерообразную форму, семена кандыма и джузгуна снабжены упругими щетинками, у саксаула – перепонками, у селина – летучками и т. п. Благодаря этому парусность плодов и семян увеличивается, и они разносятся по пустыне на большие расстояния.
Растения со- |
|
лончаков – галофи- |
|
ты безлистны или |
|
мелколистны, с |
|
толстыми членис- |
Рис. 9.141. Приспособления плодов и семян псаммофитов для переноса ветром, по |
тыми веточками и |
М. П. Петрову: |
листьями, обычно |
1 – солончаковый саксаул; 2 – джузгун; 3 – кандым; 4 – черкез Рихтера; 5 – песчаная |
акация; 6 – селин; 7 – джузгун безлистный; 8 – кумарчик широколистный |
сочными и мясис- |
|
тыми. Клеточный сок их насыщен водора- |
оригинальные жизненные формы растений |
створимыми солями – сульфатами и хлори- |
– эфемеры и эфемероиды. |
дами. Иногда содержание солей доходит до |
Эфемеры – травянистые однолетние |
45% от абсолютно сухого веса листьев. Оби- |
растения с коротким вегетационным пери- |
лие водорастворимых солей повышает ос- |
одом; они быстро, за 1 – 2 весенних месяца, |
мотическое давление клеточного сока, что |
проходят все стадии развития и, образовав |
242
семена, погибают. Неблагоприятные периоды летней жары и зимних холодов они переносят в виде семян. Эфемероиды – многолетние травянистые растения с коротким периодом вегетации. Они проходят все стадии развития и образуют семена также весной в течение 1,5 – 2 месяцев. С наступлением летней засухи наземная
часть растений высыха- Рис. 9.144. Распределение корневых систем в зарослях черного саксаула
ет, подземные же органы (Haloxylon aphyllum), по М. П. Петрову:
(корневища, луковицы, |
1 – Haloxylon aphyllum; 2 – Calligonum turkestanicum; 3 – Ephedra |
||
strobilacea; 4 – Carex physodes; 5 – эфемеры и эфемероиды |
|||
клубни и т. п.), располо- |
|||
|
|
||
женные на глубине 20 – 25 см, переходят в |
большая активность биоты приходится на |
||
стадию покоя. |
|
переходные сезоны, особенно на весну. Зи- |
|
Ранней весной пустыня покрывается гу- |
мой погода неустойчивая. Вторжение теп- |
||
стым зеленым покровом ярко цветущих трав |
лого воздуха с Иранского нагорья приводит |
||
(рис. 9.142). Благодаря тому, что раз- |
к резким повышениям температуры на юге, |
||
витие эфемеров и эфемероидов происходит |
при холодных вторжениях возможно ее по- |
||
в благоприятных гидротермических услови- |
нижение до -30° (на севере до -50°). В юж- |
||
ях, они лишены ксероморфных признаков. |
ных пустынях устойчивый снежный покров |
||
Многие из них – выходцы из северных лу- |
бывает редко. На севере устойчивый покров |
||
ãîâ. |
|
наблюдается с конца нояб-ря – декабря до |
|
В формировании растительного покро- |
марта. Мощность его невелика – 5 – 15 см. |
||
ва песчаных пустынь умеренного пояса при |
В южной подзоне в марте-апреле гос- |
||
нимает участие много разнообразных жиз- |
подствует зеленый аспект эфемеров и эфе- |
||
ненных форм (Нечаева, Василевская, Анто- |
мероидов, создаются максимальные запа- |
||
нова, 1973), благодаря чему сообщества ха- |
сы зеленой массы. Но ко второй декаде мая |
||
рактеризуются достаточно высокой видовой |
(в сухие годы – в апреле) эти растения пол- |
||
насыщенностью, сложной структурой и вы- |
ностью засыхают. В северной подзоне ве- |
||
сокой продуктивностью. Фитомасса бело- |
гетация начинается во второй половине |
||
саксаульников – около 270 ц/га, с ежегодным |
марта – начале апреля. Зацветают ранние |
||
приростом 75 ц/га; фитомасса черносаксауль- |
эфемеры, затем эфемероиды, появляется |
||
ников – 540 ц/га. |
|
листва на кустарниках и полукустарниках. |
|
В Каракумах в черно- и белосаксауло- |
Основной период вегетации – апрель - май. |
||
вой формациях четко обособляются два или |
В июне растения выглядят угнетенными; в |
||
даже три надземных яруса; соответственно |
первой декаде июня заканчивается плодо- |
||
этому ярусное распределение имеют и кор- |
ношение большинства эфемеров и эфеме- |
||
невые системы (рис. 9.143, 9.144). |
роидов, у полукустарничков перестают ра- |
||
Сезонный ритм жизненных процессов |
сти побеги и опадает часть листьев. |
||
в пустыне определяется своеобразным со- |
Лето – время длительной засухи. На |
||
четанием теплового и водного режимов, при |
юге с июня по сентябрь растительный по- |
||
котором самый теплый период оказывается |
кров выгорает. На севере в июле - августе |
||
и самым сухим, а относительно влажная |
большинство растений пребывает в состо- |
||
зима недостаточно теплой. Поэтому наи- |
янии покоя, но в сентябре начинается осен- |
||
|
|
няя вегетация. Наблюдается цветение и мак- |
|
|
|
243 симальный прирост у полукустарничков, в |
|
октябре они уже плодоносят и к концу ме-

|
|
|
|
нейших геологических |
||
|
|
|
|
эпох, другие сформиро- |
||
|
|
|
|
вались на протяжении |
||
|
|
|
|
последнего четвертич- |
||
|
|
|
|
ного периода. При этом |
||
|
|
|
|
наряду с расхождением |
||
|
|
|
|
признаков, наблюдается |
||
|
|
|
|
конвергентный путь эво- |
||
|
|
|
|
люции, когда далекие в |
||
|
|
|
|
систематическом отно- |
||
|
|
|
|
шении организмы, бла- |
||
|
|
|
|
годаря сходству условий |
||
|
|
|
|
существования приобре- |
||
|
|
|
|
тают сходный облик. Это |
||
|
|
|
|
находит отражение в |
||
|
|
|
|
сходстве строения и фун- |
||
|
|
|
|
кционирования зональ- |
||
Рис. 9.143. Густые заросли черного саксаула (Haloxylon aphyllum) |
ных типов биомов.Сво- |
|||||
еобразие природных зон |
||||||
с богатой травянистой растительностью на слабо засоленной влажной |
||||||
определяет особенности |
||||||
почве, по М. П. Петрову |
|
|
|
|||
|
|
|
|
|
||
сяца прекращают вегетировать. Во |
|
|
|
жизни человека, одно- |
||
|
|
|
|
|
||
влажные годы в это время может |
|
|
|
|
|
|
возобновиться вегетация эфемеров |
|
|
|
|
|
|
и эфемероидов. Если затем после- |
|
|
|
|
|
|
дует теплая зима, в южной подзо- |
|
|
|
|
|
|
не эфемероиды могут вегетировать |
|
|
|
|
|
|
непрерывно (как озимые) почти до |
|
|
|
|
|
|
200 дней, тогда как в сухие годы – |
|
|
|
|
|
|
лишь около 60 дней (как яровые). |
|
|
|
|
|
|
* * * |
|
|
|
|
|
|
|
Рис. 9.142. Сообщество эфемеров и эфемероидов, по Е. П. Коровину: |
|||||
Завершая обзор зональных ти- |
1 – Carex pachystylis; 2 – |
Poa bulbosa var. vivipara; 3 – Ranunculus |
||||
пов биомов суши, подчеркнем ог- |
pinnalifidus; 4 – Scaligeria alloides; 5 – Haplophyllum perforatum; |
|||||
ромную потенцию биоты к завое- |
6 – Phlomis thapsoides; |
7 – Ixiolirion tataricum; 8 |
– Gentiana |
|||
ванию жизненного пространства. olivieri; 9 – Eremostachys labiosa |
|
|
||||
На протяжении геологической ис- |
|
типные приемы ведения хозяйства и мероп- |
||||
тории эволюция биоты шла по пути дивер- |
||||||
риятия по сохранению и приумножению |
||||||
генции и экологической адаптации. По мере |
||||||
биологических ресурсов. |
Антропо- |
|||||
|
|
|||||
усиливающегося давления жизни, борьбы за |
генное воздействие на биосферу достигло в |
|
существование и естественного отбора воз- |
||
наши дни глобальных масштабов. Зональ- |
||
никали все новые формы, способные зах- |
||
ные типы биомов в естественном виде по- |
||
ватывать территории, где напряженность |
||
чти не сохранились. Проблема изменения |
||
экологических факторов находится как в ми- |
||
биосферы в результате антропогенного воз- |
||
нимальных, так и в максимальных преде- |
||
действия требует специального рассмотре- |
||
лах. В настоящее время практически все по- |
||
íèÿ. |
||
верхностные оболочки Земли представля- |
||
|
||
ют собой арену распространения жизни. |
|
|
Процессы адаптации к новым услови- |
|
|
ям жизни протекают непрерывно. На стро- |
|
|
ении одних организмов лежит печать древ- |
|
244