

2 курс / физиология / документы / Двигательная система (2)
.doc
Двигательная система.
Понятие: совокупность центральных и периферических структур, обеспечивающих деятельность опорнодвигательного аппарата.
Общий план строения ДС.
Имеет 2 отдела.
1) Сенсорный – восприятие, проведение, переработка афферентной информации, формирование двигательных программ и коррекция их по ходу выполнения.
2) Моторный отдел обеспечивает реализацию двигательных программ. Характеристика двигательных функций.
1) Поддержание позы обеспечивается путем изменения тонуса мышц и удержание сустава в нужном положении.
Активность медленных моторных единиц, устойчивых к утомлению.
2) Движения:
а) коммуникативные;
б) рабочие.
3) Выработка навыков заключается в установлении согласованной деятельности различных групп мышц.
Вначале выработки навыка вследствие иррадиации возбуждения активизируются многие центры: соматические, вегетативные. Возникают эмоции.
По мере выработки навыков путем торможения отключаются ненужные центры, движения становятся экономичными, появляется возможность их коррекции по ходу выполнения.
Эффекты двигательной системы.
I группа специфические:
а) поддержание позы путем изменения тонуса мышц за счет медленных неутомляемых моторных единиц;
б) движения → рабочие
↓
коммуникативные
Общий вид ДС
Внешний стимул → сенсорный отдел → внутренний стимул
фото ↓ проприо
ф оно
моторный отдел висцеро
оно
моторный отдел висцеро
вкусовые ↓ ↓ вестибуло
обонятельные поза движения термо
термо болевые
болевые
неспецифический эффект
Отражают связь ДС с другими системами организма:
а) моторно-висцеральные (связь с АНС);
б) психо-эмоциональные явления (связь с ЛС, корой, РФ).
ХАРАКТЕРИСТИКА СЕНСОРНОГО ОТДЕЛА ДС.
Рецепторы
- вестибуло
- тактильные
- проприо.
Афферентные пути.
I Тонкий (Голля) и клиновидный (Бурдаха) – проходит в задних рогах спинного мозга.
По волокнам тонкого пучка проводится возбуждение от нижней части тела и нижних конечностей.
По волокнам клиновидного пучка возбуждение проводится от верхней части туловища и верхних конечностей.
I нейрон – в спинальном ганглии;
II нейрон – в продолговатом мозге в соответствующих ядрах, затем после перекрестка в латеральное ядро таламуса, затем в соматосенсорную зону коры в ЗЦИ.
Скорость проведения 60 – 100м/с. Обеспечивает кожно-механическую тонко дифференцированную чувствительность, т. е. локализацию действия раздражителя, изменение его во времени.
Волокна этого пути дают коллатерали на мотонейроны и интер нейроны своего и вышележащего сегмента спинного мозга, образуя межсегментарные связи.
II Спиноталамический путь.
Латеральный – проводит болевую и температурную чувствительность.
Вентральный – тактическую чувствительность, медленнопроводящий 1 – 30м/с. Дает информацию о качественной природе раздражителей.
III Спиномозжечковый путь.
Дорзальный – не перемещаемый Флексига → информация от мышц и связок конечностей.
Вентральный путь Говерса дважды перекрещенный через продолговатый мозг и ножки мозга к коре мозжечка – от сухожильных, кожных и висцерорецепторов. Обеспечивает тонус мышц при движении и позу.
IV Вестибулярные пути к коре обеспечивают регуляцию тонуса мышц при изменении положения тела в пространстве для удержания равновесия.
Центральные структуры сенсорного отдела.
1) Кора. В каждом полушарии выделяются зоны соматической чувствительности – соматосенсорные зоны.
Первая расположена в ЗЦИ, вторая в сильвиевой борозде.
Принципы организации ССЗК:
1) Соматопическая организация, т. е. у различных рецептивных полей имеется определенная область проекции в ЗЦИ.
2) Сенсорный гомункулюс, т. е. различные рецептивные поля имеют различную площадь представительства в ЗЦИ.
Роль соматосенсорной зоны коры – интеграция и критическая оценка информации от специфических ядер таламуса. В этой же области находится и представительство висцерорецепторов.
ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА МОТОРНОГО ОТДЕЛА ДВИГАТЕЛЬНОЙ СИСТЕМЫ.
I зона – в передней центральной извилине.
II зона – в межполушарной щели.
Структурно – функциональная организация МЗК.
1) Соматотопическая организация – каждая область тела имеет свою область представительства в ПЦИ.
2) Моторный гомункулюс площадь представительства периферических отделов в ПЦИ зависит от выполняемой функции. Чем разнообразнее и точнее движения выполняет группа мышц, тем больше ее представительство в ПЦИ.
3) Наличие функциональных колонок.
Функциональное объединение вертикально расположенных нейронов.
Действуют одновременно, активизируют все мышцы, приводящие сустав в определенное положение.
Нейронная организация МЗК.
I Большие пирамидные клетки иннервируют разные мотонейроны.
Малые – тонические.
II Большие α МН
Малые γ МН
Эфферентные связи коры.
1) Пирамидный путь → к – спинальный.
↓
к – бульбарный.
Быстрое включение дыхательной системы, произвольная регуляция тонуса и движений.
2) Экстрапирамидный – коррекция двигательных программ в ходе их выполнения:
а) кортикоталамический – коррекция позы и движений
б) кортикоретикулярный → ПМ → СМ – α МН сгибателей.
↓
ВМ → СМ – γ МН разгибателей
в) к базальным ганглиям → СМ – стереотипные движения.
г) к мозжечку (через ядра моста) – коррекция движений.
д) кортикорубральные → СМ повышает тонус сгибателей, координация тонуса разных групп мышц.
е) к нижней оливе → вестибулоспинальный путь – повышает тонус разгибателей.
Роль спинного мозга в регуляции мышечного тонуса и двигательной активности
-
Для поддержания мышечного тонуса достаточно рефлекторной деятельности спинного мозга, в этом случае импульсы к мышце поступают от альфа-мотонейронов спинного мозга.
-
Двигательные рефлексы, за которые ответственен спинной мозг, осуществляются без участия сознанная, однако связь спинного мозга с головным является необходимой.
-
В процессе регуляции рефлекторной деятельности со стороны отделов головного мозга задействованы гамма-мотонейроны спинного мозга.
Роль структур головного мозга в регуляции движений и мышечного тонуса
Автоматизирование движений и их содружественность находятся под контролем базальных ганглиев.
Считается, что базальные ганглии являются высшим эфферентным центром ствола головного мозга и регулируют двигательные безусловные рефлексы, а также вегетативные реакции, сопровождающие данные рефлексы.
Предполагается также, что базальные ганглии получают информацию от ассоциативных зон коры больших полушарий, данная информация передается в таламус, куда также поступают сигналы от мозжечка. Собранная информация направляется к двигательной зоне коры больших полушарий, где формируется программа действия (целенаправленного движения), далее импульсы поступают в нижележащие отделы ЦНС, ответственные за выполнение движения.
Выделяют два типа статических рефлексов: позные (сохраняется поза) и рефлексы выпрямления (например, при переходе из состояния лежа или сидя в состояние стоя).
К статическим рефлексам относятся: лабиринтные рефлексы (возникают при изменениях положения головы в пространстве, импульсы поступают от рецепторов вестибулярного анализатора); шейные (возникают при изменениях положения головы относительно туловища, импульсы поступают от проприорецепторов шеи); выпрямительные (импульсы поступают от рецепторов кожи, вестибулярного и зрительного анализаторов).
Статокинетические рефлексы помогают сохранить равновесие при действии на организм ускорения (линейный и лифтный рефлексы), а также при повороте головы, туловища в сторону, противоположную движению (вращательный рефлекс). Статические рефлексы направлены на изменение тонуса скелетных мышц при изменении положения тела в пространстве, а также на перераспределение тонуса мышц, направленное на восстановление нормальной позы, если животное выведено из нее. В связи с этим статические рефлексы условно делят на позно-тонические и выпрямительные. Выпрямительные рефлексы проявляются в том, что животное из «непривычного», не свойственного для него положения переходит в естественное положение.
АЛЬФА И МОТОНЕЙРОНЫ
В спинном мозге располагается основной элемент всей двигательной системы мозга – альфа-мотонейроны. Аксоны этих нейронов являются единственным каналом, соединяющим нервную систему со скелетными мышцами. Возбуждение альфа-мотонейрона приводит к активации соответствующих мышечных волокон. В спинном мозге существуют два механизма, активирующих альфа-мотонейроны. Первый механизм – это прямое нисходящее влияние на альфа-мотонейрон, например, такие воздействия могут оказывать аксоны гигантских пирамидных клеток Беца, расположенных в двигательной коре. Однако активация альфа-мото-нейронов нередко осуществляется опосредованно, через вставочные нейроны (число которых в спинном мозге огромно) и гамма-мотонейроны. Последние активируют интрафузальные мышечные волокна, повышая тем самым чувствительность мышечных рецепторов, т. е. мышечных веретен к растяжению мышцы. Следствием этого является повышение потока импульсов, идущих от мышечных веретен к альфа-мотонейронам (в том числе через вставочные нейроны), что приводит к возбуждению альфа-мотонейронов и иннервируемых ими мышечных волокон. Такой механизм активации альфа-мотонейронов называется гамма-петлей. Итак, гамма-мотонейроны, по сути, выступают в роли вставочных нейронов, но с особым вариантом возбуждения, т. е. с участием периферического посредника в виде мышечных веретен. Кроме того, в спинном мозге имеются нейроны, выполняющие роль пейсмекера, т. е. автоматического генератора импульсов, возбуждающих альфа-мотонейроны. За счет такого механизма автоматически, без сигналов от супраспинальных двигательных систем могут возникнуть шагательные движения. Эти нейроны формируют так называемый центральный генератор шагания, который у здорового человека, вероятнее всего, полностью переходит под контроль супраспинальных структур (уже с первых месяцев постнатального развития).
-
Нижнетеменные области коры способствуют точной адресации команд к определенным мышцам.
-
Переднелобные зоны участвуют в программировании и регуляции произвольных движений.
-
Зоны коры, относящиеся к лимбической системе, обеспечивают эмоциональную окраску движений.
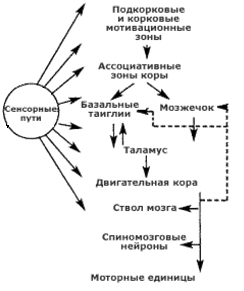
Ствол мозга, мозжечок, базальные ганглии, кора больших полушарий и их роль в регуляции движений и мышечного тонуса
Взаимосвязь отделов ЦНС, участвующих в осуществлении движений
Моторные зоны коры у приматов и человека расположены в предцентральной области, т. е. в передней центральной извилине и задних отделах верхней и средней лобных извилин. Кроме них, на медиальной поверхности коры расположена дополнительная моторная область. Электрическая стимуляция различных участков моторной коры вызывает четкие, координированные двигательные реакции, а также (при слабом раздражении) сокращения отдельных мышц противоположной половины тела.