

Molecular Biology of the Cell Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter Fourth Edition.
15. Cell Communication
According to the fossil record, sophisticated unicellular organisms resembling present-day bacteria were present on Earth for about 2.5 billion years before the first multicellular organisms appeared. One reason why multicellularity was so slow to evolve may have been related to the difficulty of developing the elaborate cell communication mechanisms that a multicellular organism needs. Its cells have to be able to communicate with one another in complex ways if they are to be able to govern their own behavior for the benefit of the organism as a whole.
These communication mechanisms depend heavily on extracellular signal molecules, which are produced by cells to signal to their neighbors or to cells further away. They also depend on elaborate systems of proteins that each cell contains to enable it to respond to a particular subset of these signals in a cell-specific way. These proteins include cell-surface receptor proteins, which bind the signal molecule, plus a variety of intracellular signaling proteins that distribute the signal to appropriate parts of the cell. Among the intracellular signaling proteins are kinases, phosphatases, GTP-binding proteins, and many other proteins with which they interact. At the end of each intracellular signaling pathway are target proteins, which are altered when the pathway is active and change the behavior of the cell. Depending on the signal's effect, these target proteins can be gene regulatory proteins, ion channels, components of a metabolic pathway, parts of the cytoskeleton, and so on (Figure 15-1).
We begin this chapter by discussing the general principles of cell communication. We then consider, in turn, each of the main families of cell-surface receptor proteins and the intracellular signaling pathways they activate. The main focus of the chapter is on animal cells, but we end by considering the special features of cell communication in plants.
 Figure
15-1. A simple intracellular
signaling pathway activated by an extracellular signal molecule.
The signal molecule binds to a receptor protein (which is usually
embedded in the plasma membrane), thereby activating an intracellular
signaling pathway that is mediated by a series of signaling proteins.
Finally, one or more of these intracellular signaling proteins
interacts with a target protein, altering the target protein so that
it helps to change the behavior of the cell.
Figure
15-1. A simple intracellular
signaling pathway activated by an extracellular signal molecule.
The signal molecule binds to a receptor protein (which is usually
embedded in the plasma membrane), thereby activating an intracellular
signaling pathway that is mediated by a series of signaling proteins.
Finally, one or more of these intracellular signaling proteins
interacts with a target protein, altering the target protein so that
it helps to change the behavior of the cell.
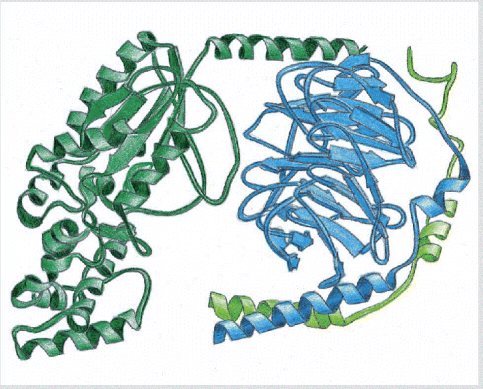 A
trimeric GTP-binding protein, or G protein. This type of G
protein functionally couples transmembrane receptors to either
enzymes or ion channels in the plasma membrane. (Based on D.G.
Lombright et al., Nature 379:311
A
trimeric GTP-binding protein, or G protein. This type of G
protein functionally couples transmembrane receptors to either
enzymes or ion channels in the plasma membrane. (Based on D.G.
Lombright et al., Nature 379:311![]() 319,
1996.)
319,
1996.)
General Principles of Cell Communication
Mechanisms enabling one cell to influence the behavior of another almost certainly existed in the world of unicellular organisms long before multicellular organisms appeared on Earth. Evidence comes from studies of present-day unicellular eucaryotes such as yeasts. Although these cells normally lead independent lives, they can communicate and influence one another's behavior in preparation for sexual mating. In the budding yeast Saccharomyces cerevisiae, for example, when a haploid individual is ready to mate, it secretes a peptide mating factor that signals cells of the opposite mating type to stop proliferating and prepare to mate (Figure 15-2). The subsequent fusion of two haploid cells of opposite mating types produces a diploid cell, which can then undergo meiosis and sporulate, generating haploid cells with new assortments of genes.
Studies of yeast mutants that are unable to mate have identified many proteins that are required in the signaling process. These proteins form a signaling network that includes cell-surface receptor proteins, GTP-binding proteins, and protein kinases, each of which has close relatives among the proteins that carry out signaling in animal cells. Through gene duplication and divergence, however, the signaling systems in animals have become much more elaborate than those in yeasts.
Extracellular Signal Molecules Bind to Specific Receptors
Yeast cells communicate with one another for mating by secreting a few kinds of small peptides. In contrast, cells in higher animals communicate by means of hundreds of kinds of signal molecules. These include proteins, small peptides, amino acids, nucleotides, steroids, retinoids, fatty acid derivatives, and even dissolved gases such as nitric oxide and carbon monoxide. Most of these signal molecules are secreted from the signaling cell into the extracellular space by exocytosis (discussed in Chapter 13). Others are released by diffusion through the plasma membrane, and some are exposed to the extracellular space while remaining tightly bound to the signaling cell's surface.
Regardless of the nature of the signal, the target cell responds by means of a specific protein called a receptor, which specifically binds the signal molecule and then initiates a response in the target cell. The extracellular signal molecules often act at very low concentrations (typically 10-8 M), and the receptors that recognize them usually bind them with high affinity (affinity constant Ka 108 liters/mole; see Figure 3-44). In most cases, these receptors are transmembrane proteins on the target cell surface. When they bind an extracellular signal molecule (a ligand), they become activated and generate a cascade of intracellular signals that alter the behavior of the cell. In other cases, the receptors are inside the target cell, and the signal molecule has to enter the cell to activate them: this requires that the signal molecules be sufficiently small and hydrophobic to diffuse across the plasma membrane (Figure 15-3).
Extracellular Signal Molecules Can Act Over Either Short or Long Distances
Many signal molecules remain bound to the surface of the signaling cell and influence only cells that contact it (Figure 15-4A). Such contact-dependent signaling is especially important during development and in immune responses. In most cases, however, signal molecules are secreted. The secreted molecules may be carried far afield to act on distant targets, or they may act as local mediators, affecting only cells in the immediate environment of the signaling cell. This latter process is called paracrine signaling (Figure 15-4B). For paracrine signals to be delivered only to their proper target cells, the secreted molecules must not be allowed to diffuse too far; for this reason they are often rapidly taken up by neighboring target cells, destroyed by extracellular enzymes, or immobilized by the extracellular matrix.
For a large, complex multicellular organism, short-range signaling is not sufficient on its own to coordinate the behavior of its cells. In these organisms, sets of specialized cells have evolved with a specific role in communication between widely separate parts of the body. The most sophisticated of these are nerve cells, or neurons, which typically extend long processes (axons) that enable them to contact target cells far away. When activated by signals from the environment or from other nerve cells, a neuron sends electrical impulses (action potentials) rapidly along its axon; when such an impulse reaches the end of the axon, it causes the nerve terminals located there to secrete a chemical signal called a neurotransmitter. These signals are secreted at specialized cell junctions called chemical synapses, which are designed to ensure that the neurotransmitter is delivered specifically to the postsynaptic target cell (Figure 15-4C). The details of this synaptic signaling process are discussed in Chapter 11.
A second type of specialized signaling cell that controls the behavior of the organism as a whole is an endocrine cell. These cells secrete their signal molecules, called hormones, into the bloodstream, which carries the signal to target cells distributed widely throughout the body (Figure 15-4D).
The mechanisms that allow endocrine cells and nerve cells to coordinate cell behavior in animals are compared in Figure 15-5. Because endocrine signaling relies on diffusion and blood flow, it is relatively slow. Synaptic signaling, by contrast, can be much faster, as well as more precise. Nerve cells can transmit information over long distances by electrical impulses that travel at rates of up to 100 meters per second; once released from a nerve terminal, a neurotransmitter has to diffuse less than 100 nm to the target cell, a process that takes less than a millisecond. Another difference between endocrine and synaptic signaling is that, whereas hormones are greatly diluted in the bloodstream and interstitial fluid and therefore must be able to act at very low concentrations (typically < 10-8 M), neurotransmitters are diluted much less and can achieve high local concentrations. The concentration of acetylcholine in the synaptic cleft of an active neuromuscular junction, for example, is about 5 × 10-4 M. Correspondingly, neurotransmitter receptors have a relatively low affinity for their ligand, which means that the neurotransmitter can dissociate rapidly from the receptor to terminate a response. Moreover, after its release from a nerve terminal, a neurotransmitter is quickly removed from the synaptic cleft, either by specific hydrolytic enzymes that destroy it or by specific membrane transport proteins that pump it back into either the nerve terminal or neighboring glial cells. Thus, synaptic signaling is much more precise than endocrine signaling, both in time and in space.
The speed of a response to an extracellular signal depends not only on the mechanism of signal delivery, but also on the nature of the response in the target cell. Where the response requires only changes in proteins already present in the cell, it can occur in seconds or even milliseconds. When the response involves changes in gene expression and the synthesis of new proteins, however, it usually requires hours, irrespective of the mode of signal delivery.
Autocrine Signaling Can Coordinate Decisions by Groups of Identical Cells
All of the forms of signaling discussed so far allow one cell to influence another. Often, the signaling cell and target are different cell types. Cells, however, can also send signals to other cells of the same type, as well as to themselves. In such autocrine signaling, a cell secretes signal molecules that can bind back to its own receptors. During development, for example, once a cell has been directed along a particular pathway of differentiation, it may begin to secrete autocrine signals to itself that reinforce this developmental decision.
Autocrine signaling is most effective when performed simultaneously by neighboring cells of the same type, and it is likely to be used to encourage groups of identical cells to make the same developmental decisions. Thus, autocrine signaling is thought to be one possible mechanism underlying the "community effect" that is observed in early development, during which a group of identical cells can respond to a differentiation-inducing signal but a single isolated cell of the same type cannot (Figure 15-6).
Unfortunately, cancer cells often use autocrine signaling to overcome the normal controls on cell proliferation and survival that we discuss later. By secreting signals that act back on the cell's own receptors, cancer cells can stimulate their own survival and proliferation and thereby survive and proliferate in places where normal cells of the same type could not. How this dangerous perturbation of normal cell behavior comes about is discussed in Chapter 23.
Gap Junctions Allow Signaling Information to Be Shared by Neighboring Cells
Another way to coordinate the activities of neighboring cells is through gap junctions. These are specialized cell-cell junctions that can form between closely apposed plasma membranes and directly connect the cytoplasms of the joined cells via narrow water-filled channels (see Figure 19-15). The channels allow the exchange of small intracellular signaling molecules (intracellular mediators), such as Ca2+ and cyclic AMP (discussed later), but not of macromolecules, such as proteins or nucleic acids. Thus, cells connected by gap junctions can communicate with each other directly, without having to surmount the barrier presented by the intervening plasma membranes (Figure 15-7).
As discussed in Chapter 19, the pattern of gap-junction connections in a tissue can be revealed either electrically, with intracellular electrodes, or visually, after the microinjection of small water-soluble dyes. Studies of this kind indicate that the cells in a developing embryo make and break gap-junction connections in specific and interesting patterns, strongly suggesting that these junctions have an important role in the signaling processes that occur between these cells. Mice and humans that are deficient in one particular gap-junction protein (connexin 43), for example, have severe defects in heart development. Like the autocrine signaling described above, gap-junction communication helps adjacent cells of a similar type to coordinate their behavior. It is still not known, however, which particular small molecules are important as carriers of signals through gap junctions, and the specific functions of gap-junction communication in animal development remain uncertain.
Each Cell Is Programmed to Respond to Specific Combinations of Extracellular Signal Molecules
A typical cell in a multicellular organism is exposed to hundreds of different signals in its environment. These signals can be soluble, bound to the extracellular matrix, or bound to the surface of a neighboring cell, and they can act in many millions of combinations. The cell must respond to this babel of signals selectively, according to its own specific character, which it has acquired through progressive cell specialization in the course of development. A cell may be programmed to respond to one combination of signals by differentiating, to another combination by multiplying, and to yet another by performing some specialized function such as contraction or secretion.
Most of the cells in a complex animal are also programmed to depend on a specific combination of signals simply to survive. When deprived of these signals (in a culture dish, for example), a cell activates a suicide program and kills itselfa process called programmed cell death, or apoptosis (Figure 15-8). Because different types of cells require different combinations of survival signals, each cell type is restricted to different environments in the body. The ability to undergo apoptosis is a fundamental property of animal cells, and it is discussed in Chapter 17.
In principle, the hundreds of signal molecules that animals make can be used to create an almost unlimited number of signaling combinations. The use of these combinations to control cell behavior enables an animal to control its cells in highly specific ways by using a limited diversity of signal molecules.
Different Cells Can Respond Differently to the Same Extracellular Signal Molecule
The specific way in which a cell reacts to its environment varies. It varies according to the set of receptor proteins the cell possesses, which determines the particular subset of signals it can respond to, and it varies according to the intracellular machinery by which the cell integrates and interprets the signals it receives (see Figure 15-1). Thus, a single signal molecule often has different effects on different target cells. The neurotransmitter acetylcholine, for example, stimulates the contraction of skeletal muscle cells, but it decreases the rate and force of contraction in heart muscle cells. This is because the acetylcholine receptor proteins on skeletal muscle cells are different from those on heart muscle cells. But receptor differences are not always the explanation for the different effects. In many cases, the same signal molecule binds to identical receptor proteins yet produces very different responses in different types of target cells, reflecting differences in the internal machinery to which the receptors are coupled (Figure 15-9).
The Concentration of a Molecule Can Be Adjusted Quickly Only If the Lifetime of the Molecule Is Short
It is natural to think of signaling systems in terms of the changes produced when a signal is delivered. But it is just as important to consider what happens when a signal is withdrawn. During development, transient signals often produce lasting effects: they can trigger a change in the cell's development that persists indefinitely, through cell memory mechanisms such as those discussed in Chapters 7 and 21. In most cases in adult tissues, however, the response fades when a signal ceases. The effect is transitory because the signal exerts its effects by altering a set of molecules that are unstable, undergoing continual turnover. Thus, once the signal is shut off, the replacement of the old molecules by new ones wipes out all traces of its action. It follows that the speed with which a cell responds to signal removal depends on the rate of destruction, or turnover, of the molecules the signal affects.
It is also true, although much less obvious, that this turnover rate also determines the promptness of the response when a signal is turned on. Consider, for example, two intracellular signaling molecules X and Y, both of which are normally maintained at a concentration of 1000 molecules per cell. Molecule Y is synthesized and degraded at a rate of 100 molecules per second, with each molecule having an average lifetime of 10 seconds. Molecule X has a turnover rate that is 10 times slower than that of Y: it is both synthesized and degraded at a rate of 10 molecules per second, so that each molecule has an average lifetime in the cell of 100 seconds. If a signal acting on the cell boosts the rates of synthesis of both X and Y tenfold without any change in the molecular lifetimes, at the end of 1 second the concentration of Y will have increased by nearly 900 molecules per cell (10 × 100 - 100), while the concentration of X will have increased by only 90 molecules per cell. In fact, after a molecule's synthesis rate has been either increased or decreased abruptly, the time required for the molecule to shift halfway from its old to its new equilibrium concentration is equal to its normal half-lifethat is, equal to the time that would be required for its concentration to fall by half if all synthesis were stopped (Figure 15-10).
The same principles apply to proteins and small molecules, and to molecules in the extracellular space and inside cells. Many intracellular proteins have short half-lives, some surviving for less than 10 minutes. In most cases, these are proteins with key regulatory roles, whose concentrations are rapidly regulated in the cell by changes in their rates of synthesis. Likewise, any covalent modifications of proteins that occur as part of a rapid signaling processmost commonly, the addition of a phosphate group to an amino acid side chainmust be continuously removed at a rapid rate to make rapid signaling possible.
We shall discuss some of these molecular events in detail later for signaling pathways that operate via cell-surface receptors. But the principles apply quite generally, as the next example illustrates.
Nitric Oxide Gas Signals by Binding Directly to an Enzyme Inside the Target Cell
Although most extracellular signals are hydrophilic molecules that bind to receptors on the surface of the target cell, some signal molecules are hydrophobic enough and/or small enough to pass readily across the target-cell plasma membrane. Once inside, they directly regulate the activity of a specific intracellular protein. An important and remarkable example is the gas nitric oxide (NO), which acts as a signal molecule in both animals and plants. In mammals, one of its functions is to regulate smooth muscle contraction. Acetylcholine, for example, is released by autonomic nerves in the walls of a blood vessel, and it causes smooth muscle cells in the vessel wall to relax. The acetylcholine acts indirectly by inducing the nearby endothelial cells to make and release NO, which then signals the underlying smooth muscle cells to relax. This effect of NO on blood vessels provides an explanation for the mechanism of action of nitroglycerine, which has been used for about 100 years to treat patients with angina (pain resulting from inadequate blood flow to the heart muscle). The nitroglycerine is converted to NO, which relaxes blood vessels. This reduces the workload on the heart and, as a consequence, it reduces the oxygen requirement of the heart muscle.
Many types of nerve cells use NO gas to signal to their neighbors. The NO released by autonomic nerves in the penis, for example, causes the local blood vessel dilation that is responsible for penile erection. NO is also produced as a local mediator by activated macrophages and neutrophils to help them to kill invading microorganisms. In plants, NO is involved in the defensive responses to injury or infection.
NO gas is made by the deamination of the amino acid arginine, catalyzed by the enzyme NO synthase. Because it passes readily across membranes, dissolved NO rapidly diffuses out of the cell where it is produced and into neighboring cells. It acts only locally because it has a short half-lifeabout 510 secondsin the extracellular space before it is converted to nitrates and nitrites by oxygen and water. In many target cells, including endothelial cells, NO binds to iron in the active site of the enzyme guanylyl cyclase, stimulating this enzyme to produce the small intracellular mediator cyclic GMP, which we discuss later (Figure 15-11). The effects of NO can occur within seconds, because the normal rate of turnover of cyclic GMP is high: a rapid degradation to GMP by a phosphodiesterase constantly balances the production of cyclic GMP from GTP by guanylyl cyclase. The drug Viagra inhibits this cyclic GMP phosphodiesterase in the penis, thereby increasing the amount of time that cyclic GMP levels remain elevated after NO production is induced by local nerve terminals. The cyclic GMP, in turn, keeps blood vessels relaxed and the penis erect.
Carbon monoxide (CO) is another gas that is used as an intercellular signal. It can act in the same way as NO, by stimulating guanylyl cyclase. These gases are not the only signal molecules that can pass directly across the target-cell plasma membrane. A group of small, hydrophobic, nongaseous hormones and local mediators also enter target cells in this way. But instead of binding to enzymes, they bind to intracellular receptor proteins that directly regulate gene transcription, as we discuss next.
Nuclear Receptors Are Ligand-activated Gene Regulatory Proteins
A number of small hydrophobic signal molecules diffuse directly across the plasma membrane of target cells and bind to intracellular receptor proteins. These signal molecules include steroid hormones, thyroid hormones, retinoids, and vitamin D. Although they differ greatly from one another in both chemical structure (Figure 15-12) and function, they all act by a similar mechanism. When these signal molecules bind to their receptor proteins, they activate the receptors, which bind to DNA to regulate the transcription of specific genes. The receptors are all structurally related, being part of the nuclear receptor superfamily. This very large superfamily also includes some receptor proteins that are activated by intracellular metabolites rather than by secreted signal molecules. Many family members have been identified by DNA sequencing only, and their ligand is not yet known; these proteins are therefore referred to as orphan nuclear receptors. The importance of such nuclear receptors in some animals is indicated by the fact that 12% of the genes in the nematode C. elegans code for them, although there are fewer than 50 in humans (see Figure 7-114).
Steroid hormoneswhich include cortisol, the steroid sex hormones, vitamin D (in vertebrates), and the moulting hormone ecdysone (in insects)are all made from cholesterol. Cortisol is produced in the cortex of the adrenal glands and influences the metabolism of many types of cells. The steroid sex hormones are made in the testes and ovaries, and they are responsible for the secondary sex characteristics that distinguish males from females. Vitamin D is synthesized in the skin in response to sunlight; after it has been converted to its active form in the liver or kidneys, it regulates Ca2+ metabolism, promoting Ca2+ uptake in the gut and reducing its excretion in the kidneys. The thyroid hormones, which are made from the amino acid tyrosine, act to increase the metabolic rate in a wide variety of cell types, while the retinoids, such as retinoic acid, are made from vitamin A and have important roles as local mediators in vertebrate development. Although all of these signal molecules are relatively insoluble in water, they are made soluble for transport in the bloodstream and other extracellular fluids by binding to specific carrier proteins, from which they dissociate before entering a target cell (see Figure 15-3).
Beside a fundamental difference in the way they signal their target cells, most water-insoluble signal molecules differ from water-soluble ones in the length of time they persist in the bloodstream or tissue fluids. Most water-soluble hormones are removed and/or broken down within minutes of entering the blood, and local mediators and neurotransmitters are removed from the extracellular space even fasterwithin seconds or milliseconds. Steroid hormones, by contrast, persist in the blood for hours and thyroid hormones for days. Consequently, water-soluble signal molecules usually mediate responses of short duration, whereas water-insoluble ones tend to mediate responses that are longer lasting.
The intracellular receptors for the steroid and thyroid hormones, retinoids, and vitamin D all bind to specific DNA sequences adjacent to the genes the ligand regulates. Some receptors, such as those for cortisol, are located primarily in the cytosol and enter the nucleus after ligand binding; others, such as the thyroid and retinoid receptors, are bound to DNA in the nucleus even in the absence of ligand. In either case, the inactive receptors are bound to inhibitory protein complexes, and ligand binding alters the conformation of the receptor protein, causing the inhibitory complex to dissociate. The ligand binding also causes the receptor to bind to coactivator proteins that induce gene transcription (Figure 15-13). The transcriptional response usually takes place in successive steps: the direct activation of a small number of specific genes occurs within about 30 minutes and constitutes the primary response; the protein products of these genes in turn activate other genes to produce a delayed, secondary response; and so on. In this way, a simple hormonal trigger can cause a very complex change in the pattern of gene expression (Figure 15-14).
The responses to steroid and thyroid hormones, vitamin D, and retinoids, like responses to extracellular signals in general, are determined as much by the nature of the target cell as by the nature of the signal molecule. Many types of cells have the identical intracellular receptor, but the set of genes that the receptor regulates is different in each cell type. This is because more than one type of gene regulatory protein generally must bind to a eucaryotic gene to activate its transcription. An intracellular receptor can therefore activate a gene only if there is the right combination of other gene regulatory proteins, and many of these are cell-type specific. Thus, each of these hormones induces a characteristic set of responses in an animal for two reasons. First, only certain types of cells have receptors for it. Second, each of these cell types contains a different combination of other cell-type-specific gene regulatory proteins that collaborate with the activated receptor to influence the transcription of specific sets of genes.
The molecular details of how nuclear receptors and other gene regulatory proteins control specific gene transcription are discussed in Chapter 7.
The Three Largest Classes of Cell-Surface Receptor Proteins Are Ion-Channel-linked, G-Protein-linked, and Enzyme-linked Receptors
As mentioned previously, all water-soluble signal molecules (including neurotransmitters and all signal proteins) bind to specific receptor proteins on the surface of the target cells that they influence. These cell-surface receptor proteins act as signal transducers. They convert an extracellular ligand-binding event into intracellular signals that alter the behavior of the target cell.
Most cell-surface receptor proteins belong to one of three classes, defined by the transduction mechanism they use. Ion-channel-linked receptors, also known as transmitter-gated ion channels or ionotropic receptors, are involved in rapid synaptic signaling between electrically excitable cells (Figure 15-15A). This type of signaling is mediated by a small number of neurotransmitters that transiently open or close an ion channel formed by the protein to which they bind, briefly changing the ion permeability of the plasma membrane and thereby the excitability of the postsynaptic cell. The ion-channel-linked receptors belong to a large family of homologous, multipass transmembrane proteins. Because they are discussed in detail in Chapter 11, we shall not consider them further here.
G-protein-linked receptors act indirectly to regulate the activity of a separate plasma-membrane-bound target protein, which can be either an enzyme or an ion channel. The interaction between the receptor and this target protein is mediated by a third protein, called a trimeric GTP-binding protein (G protein) (Figure 15-15B). The activation of the target protein can change the concentration of one or more intracellular mediators (if the target protein is an enzyme), or it can change the ion permeability of the plasma membrane (if the target protein is an ion channel). The intracellular mediators affected act in turn to alter the behavior of yet other signaling proteins in the cell. All of the G-protein-linked receptors belong to a large family of homologous, seven-pass transmembrane proteins.
Enzyme-linked receptors, when activated, either function directly as enzymes or are directly associated with enzymes that they activate (Figure 15-15C). They are formed by single-pass transmembrane proteins that have their ligand-binding site outside the cell and their catalytic or enzyme-binding site inside. Enzyme-linked receptors are heterogeneous in structure compared with the other two classes. The great majority, however, are protein kinases, or are associated with protein kinases, and ligand binding to them causes the phosphorylation of specific sets of proteins in the target cell.
There are some cell-surface receptors that do not fit into any of the above classes. Some of these depend on intracellular proteolytic events to signal the cell, and we discuss them only after we explain in detail how G-protein-linked receptors and enzyme-linked receptors operate. We start with some general principles of signaling via cell-surface receptors.
Most Activated Cell-Surface Receptors Relay Signals Via Small Molecules and a Network of Intracellular Signaling Proteins
Signals received at the surface of a cell by either G-protein-linked or enzyme-linked receptors are relayed into the cell interior by a combination of small and large intracellular signaling molecules. The resulting chain of intracellular signaling events ultimately alters target proteins, and these altered target proteins are responsible for modifying the behavior of the cell (see Figure 15-1).
The small intracellular signaling molecules are called small intracellular mediators, or second messengers (the "first messengers" being the extracellular signals). They are generated in large numbers in response to receptor activation and rapidly diffuse away from their source, broadcasting the signal to other parts of the cell. Some, such as cyclic AMP and Ca2+, are water-soluble and diffuse in the cytosol, while others, such as diacylglycerol, are lipid-soluble and diffuse in the plane of the plasma membrane. In either case, they pass the signal on by binding to and altering the behavior of selected signaling proteins or target proteins.
The large intracellular signaling molecules are intracellular signaling proteins. Many of these relay the signal into the cell by either activating the next signaling protein in the chain or generating small intracellular mediators. These proteins can be classified according to their particular function, although many fall into more than one category (Figure 15-16):
1. Relay proteins simply pass the message to the next signaling component in the chain.
2. Messenger proteins carry the signal from one part of the cell to another, such as from the cytosol to the nucleus.
3. Adaptor proteins link one signaling protein to another, without themselves conveying a signal.
4. Amplifier proteins, which are usually either enzymes or ion channels, greatly increase the signal they receive, either by producing large amounts of small intracellular mediators or by activating large numbers of downstream intracellular signaling proteins. When there are multiple amplification steps in a relay chain, the chain is often referred to as a signaling cascade.
5. Transducer proteins convert the signal into a different form. The enzyme that makes cyclic AMP is an example: it both converts the signal and amplifies it, thus acting as both a transducer and an amplifier.
6. Bifurcation proteins spread the signal from one signaling pathway to another.
7. Integrator proteins receive signals from two or more signaling pathways and integrate them before relaying a signal onward.
8. Latent gene regulatory proteins are activated at the cell surface by activated receptors and then migrate to the nucleus to stimulate gene transcription.
As shown in blue in Figure 15-16, other types of intracellular proteins also have important roles in intracellular signaling. Modulator proteins modify the activity of intracellular signaling proteins and thereby regulate the strength of signaling along the pathway. Anchoring proteins maintain specific signaling proteins at a precise location in the cell by tethering them to a membrane or the cytoskeleton. Scaffold proteins are adaptor and/or anchoring proteins that bind multiple signaling proteins together in a functional complex and often hold them at a specific location.
Some Intracellular Signaling Proteins Act as Molecular Switches
Many intracellular signaling proteins behave like molecular switches: on receipt of a signal they switch from an inactive to an active state, until another process switches them off. As we discussed earlier, the switching off is just as important as the switching on. If a signaling pathway is to recover after transmitting a signal so that it can be ready to transmit another, every activated molecule in the pathway must be returned to its original inactivated state.
The molecular switches fall into two main classes that operate in different ways, although in both cases it is the gain or loss of phosphate groups that determines whether the protein is active or inactive. The largest class consists of proteins that are activated or inactivated by phosphorylation (discussed in Chapter 3). For these proteins, the switch is thrown in one direction by a protein kinase, which adds one or more phosphate groups to the signaling protein, and in the other direction by a protein phosphatase, which removes the phosphate groups from the protein (Figure 15-17A). It is estimated that one-third of the proteins in a eucaryotic cell are phosphorylated at any given time.
Many of the signaling proteins controlled by phosphorylation are themselves protein kinases, and these are often organized into phosphorylation cascades. One protein kinase, activated by phosphorylation, phosphorylates the next protein kinase in the sequence, and so on, relaying the signal onward and, in the process, amplifying it and sometimes spreading it to other signaling pathways. Two main types of protein kinases operate as intracellular signaling proteins. The great majority are serine/threonine kinases, which phosphorylate proteins on serines and (less often) threonines. Others are tyrosine kinases, which phosphorylate proteins on tyrosines. An occasional kinase can do both. Genome sequencing reveals that about 2% of our genes encode protein kinases, and it is thought that hundreds of distinct types of protein kinases are present in a typical mammalian cell.
The other main class of molecular switches involved in signaling are GTP-binding proteins (discussed in Chapter 3). These switch between an active state when GTP is bound and an inactive state when GDP is bound. Once activated, they have intrinsic GTPase activity and shut themselves off by hydrolyzing their bound GTP to GDP (Figure 15-17B). There are two major types of GTP-binding proteinslarge trimeric GTP-binding proteins (also called G proteins), which relay the signals from G-protein-linked receptors (see Figure 15-15B), and small monomeric GTPases (also called monomeric GTP-binding proteins). The latter also help to relay intracellular signals, but in addition they are involved in regulating vesicular traffic and many other processes in eucaryotic cells.
As discussed earlier, complex cell behaviors, such as cell survival and cell proliferation, are generally stimulated by specific combinations of extracellular signals rather than by a single signal acting alone (see Figure 15-8). The cell therefore has to integrate the information coming from separate signals so as to make an appropriate responseto live or die, to divide or not, and so on. This integration usually depends on integrator proteins (see Figure 15-16), which are equivalent to the microprocessors in a computer: they require multiple signal inputs to produce an output that causes the desired biological effect. Two examples that show how such integrator proteins can operate are illustrated in Figure 15-18.